
广东南澳海水养殖区中小型浮游动物群落结构特征
|
王庆(1981—), 男, 安徽省安庆市人, 副研究员, 从事浮游动物生态学研究。email: wq2010@jnu.edu.cn |
Editor: 林强
收稿日期: 2024-01-20
修回日期: 2024-02-26
网络出版日期: 2024-02-29
基金资助
国家自然科学基金(32271684)
国家自然科学基金(41503072)
粤东上升流区海洋生态系统综合观测研究站2023年开放基金项目(CUE202303)
Community structure of meso- and micro-zooplankton in the mariculture area in the Nan’ao Island, Guangdong Province
Editor: LIN Qiang
Received date: 2024-01-20
Revised date: 2024-02-26
Online published: 2024-02-29
Supported by
National Natural Science Foundation of China(32271684)
National Natural Science Foundation of China(41503072)
Foundation of Guangdong Provincial Observation and Research Station for Coastal Upwelling Ecosystem(CUE202303)
海水养殖对近海环境质量具有重要影响。为调查广东汕头南澳不同类型海水养殖活动对中小型浮游动物群落结构的影响, 于2016年4月至6月在大型海藻龙须菜收获前后, 高频次调查南澳深澳湾鱼类养殖区、龙须菜栽培区、鲍养殖区和鲍藻复合养殖区的浮游动物群落结构特征。调查期间, 浮游动物共检出44种, 包括浮游幼虫8种、原生动物6种、轮虫1种、枝角类2种和桡足类27种。中小型浮游动物在不同类型调查区域的丰度变化范围为4~392ind.·L-1, 其中鲍养殖区丰度最高(P<0.05)。浮游动物优势种为钟状网纹虫(Favella campanula)、小拟哲水蚤(Paracalanus parvus)、小毛猛水蚤(Microsetella norvegica)和拟长腹剑水蚤(Oithona similis)。体长频度分析显示, 98.74%的浮游动物体长小于1mm, 主要为原生动物、浮游幼虫和无节幼体。龙须菜收获后, 原龙须菜栽培区的浮游动物群落结构变化与对照海域差异不显著。冗余分析表明, 总磷、活性磷酸盐、盐度和溶解氧等环境因子显著影响浮游动物群落结构。调查结果表明, 海水养殖设施形成微生境, 使中小型浮游动物丰度增加, 导致浮游动物群落结构小型化; 大型海藻龙须菜的规模栽培能增加浮游动物群落结构的稳定性。
王庆 , 任辉 , 柯志新 . 广东南澳海水养殖区中小型浮游动物群落结构特征[J]. 热带海洋学报, 2024 , 43(6) : 92 -103 . DOI: 10.11978/2024025
Mariculture has an important influence on the quality of coastal environment. To investigate the effects of mariculture on meso- and micro-zooplankton community structure, sampling was conducted in the different mariculture areas (fish cultivation area, Gracilaria lemaneiformis cultivation area, abalone cultivation area and abalone-G. lemaneiformis integrated cultivation area) and natural sea area in the Shen’ao Bay of the Nan’ao Island, Shantou City from April to June, 2016. During the surveys, 44 species of zooplankton, including 8 species of planktonic larvae, 6 protozoan, 1 rotifer, 2 cladocerans and 27 copepods were identified in the Shen’ao Bay. The zooplankton abundance ranged from 4 to 392 ind.·L-1, which was higher in mariculture area and most abundant in abalone cultivation area (P<0.05). The dominant species of zooplankton were Favella campanula, Paracalanus parvus, Microsetella norvegica and Oithona similis. Size-frequency distributions of zooplankton indicated that mariculture had a strong influence on the zooplankton body size. The body size of zooplankton ranged from 0.06 to 2.18 mm and most of them (98.74%) were under 1 mm with significant fluctuation, mainly protozoa, nauplius and planktonic larvae. After G. lemaneiformis harvest, there was no significant difference in changes of zooplankton community structure between the former G. lemaneiformis cultivation area and the natural sea area. Redundancy analysis indicated that total phosphorus, $\mathrm{PO}_{4}^{3-}$, salinity and dissolved oxygen were the primary environmental factors. Results showed mariculture facilities form microhabitats which increase meso- and micro-zooplankton abundance, and zooplankton community structure becomes smaller; the large-scale cultivation of seaweed G. lemaneiformis has increased the stability of zooplankton community.
Key words: zooplankton; mariculture; integrated aquaculture; seaweed; biodiversity
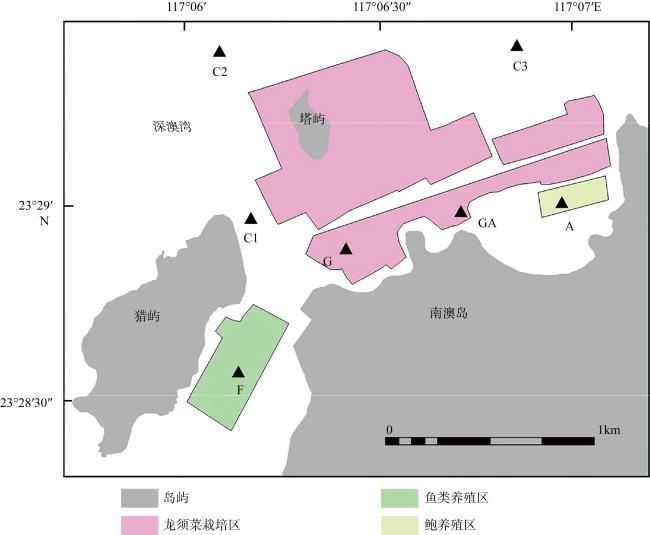
图1 汕头深澳湾海域采样站位图该图基于国家测绘地理信息局标准地图服务网站下载的审图号为粤S(2018)080号的标准地图制作。A: 鲍养殖区; C: 对照区(C1、C2和C3为对照区的三个采样点); F: 鱼类养殖区; GA: 贝藻复合养殖区; G: 龙须菜栽培区 Fig. 1 Sampling sites in the Shen’ao Bay, Nan’ao Island. A: abalone mariculture area; C: nature sea area; F: fish mariculture area; GA: G. lemaneiformis-abalone integrated mariculture area; G: G. lemaneiformis cultivation area |
图2 南澳深澳湾海域不同类型调查区域水体总氮总磷和叶绿素a时空变化A: 鲍养殖区; C: 对照区; F: 鱼类养殖区; GA: 贝藻复合养殖区; G: 龙须菜栽培区 Fig. 2 Spatial-temporal changes in total nitrogen, total phosphorus and chlorophyll a in different areas of the Shen’ao Bay, Nan’ao Island. A: Abalone mariculture area; C: nature sea area; F: fish mariculture area; GA: G. lemaneiformis-abalone integrated mariculture area; G: G. lemaneiformis cultivation area |
表1 南澳深澳湾海域不同类型调查区域浮游动物种类名录Tab. 1 Species composition of zooplankton in different areas of the Shen’ao Bay, Nan’ao Island |
| 门 | 目 | 种 |
|---|---|---|
| 纤毛门Ciliophora | 环毛目Choreotrichida | 钟状网纹虫Favella campanula |
| 砂壳目Tintinnida | 管状拟铃虫Tintinnopsis tubulosa | |
| 妥肯丁拟铃虫Tintinnopsis tocantininsis | ||
| 根状拟铃虫Tintinnopsis radix | ||
| 大拟铃虫Tintinnopsis ampla | ||
| 粘孢子门Myzozoa | 夜光藻目Noctilucales | 夜光虫Noctiluca scintillans |
| 轮虫动物门Rotifera | 游泳目Ploima | 疣毛轮虫Synchaeta sp. |
| 节肢动物门Arthropoda | 钩足目Onychopoda | 肥胖三角溞Pseudevadne tergestina |
| 栉足目Ctenopoda | 鸟喙尖头溞Penilia avirostris | |
| 哲水蚤目Calanoida | 长纺锤水蚤Acartia longiremis | |
| 驼背隆哲水蚤Acrocalanus gibber | ||
| 中华哲水蚤Calanus sinicus | ||
| 双刺平头水蚤Candacia bipinnata | ||
| 微刺哲水蚤Canthocalanus pauper | ||
| 腹针胸刺水蚤Centropages abdominalis | ||
| 达氏筛哲水蚤Cosmocalanus darwinii | ||
| 精致真刺水蚤 Euchaeta concinna | ||
| 针刺拟哲水蚤Paracalanus aculeatus | ||
| 瘦拟哲水蚤Paracalanus gracilis | ||
| 小拟哲水蚤Paracalanus parvus | ||
| 锥形宽水蚤Temora turbinata | ||
| 强额孔雀哲水蚤Parvocalanus crassirostris | ||
| 钳形歪水蚤Tortanus forcipatus | ||
| 节肢动物门Arthropoda | 剑水蚤目Cyclopoida | 平大眼剑水蚤Corycaeus dahli |
| 短大眼剑水蚤Corycaeus giesbrechti | ||
| 坚长腹剑水蚤 Dioithona rigida | ||
| 拟长腹剑水蚤Oithona similis | ||
| 小长腹剑水蚤Oithona nana | ||
| 简长腹剑水蚤Oithona simplex | ||
| 丽隆剑水蚤Oncaea venusta | ||
| 等刺隆水蚤Oncaea mediterranea | ||
| 猛水蚤目Harpacticoida | 硬鳞暴猛水蚤Clytemnestra scutellata | |
| 尖额谐猛水蚤Euterpina acutifrons | ||
| 小毛猛水蚤Microsetella norvegica | ||
| 虎斑猛水蚤Tigriopus fulvus | ||
| 猛水蚤目Harpacticoida未定种 | ||
| 端足目Amphipoda | 端足目Amphipoda未定种 | |
| 钩虾幼体Gammarid larva | ||
| 磷虾目Euphausiacea | 磷虾幼体 Euphausiidae larva | |
| 蔓足类六肢幼体Cirripedia larva | ||
| 环节动物门Annelida | 多毛类浮游幼虫Polychaeta larva | |
| 毛颚动物门Chaetognatha | 无横肌目Aphragmophora | 箭虫属未定种Sagitta sp. |
| 刺胞动物门Cnidaria | 管水母目Siphonophorae | 双生水母科Diphyidae未定种 |
| 脊索动物门Chordata | 有尾目Copelata | 住囊虫属未定种Oikopleura sp. |
图3 南澳深澳湾海域不同类型调查区域浮游动物丰度的时空变化A: 鲍养殖区; C: 对照区; F: 鱼类养殖区; GA: 贝藻复合养殖区; G: 龙须菜栽培区 Fig. 3 Spatialtemporal variations in zooplankton abundance in different areas of the Shen’ao Bay, Nan’ao Island. A: abalone mariculture area; C: nature sea area; F: fish mariculture area; GA: G. lemaneiformis-abalone integrated mariculture area; G: G. lemaneiformis cultivation area |
图4 南澳深澳湾海域不同类型调查区域浮游动物丰度及其百分比组成的时空变化A: 鲍养殖区; C: 对照区; F: 鱼类养殖区; GA: 贝藻复合养殖区; G: 龙须菜栽培区 Fig. 4 Spatial-temporal changes in zooplankton abundance and its percentage composition in different areas of the Shen’ao Bay, Nan’ao Island. A: abalone mariculture area; C: natural sea area; F: fish mariculture area; GA: G. lemaneiformis-abalone integrated mariculture area; G: G. lemaneiformis cultivation area |
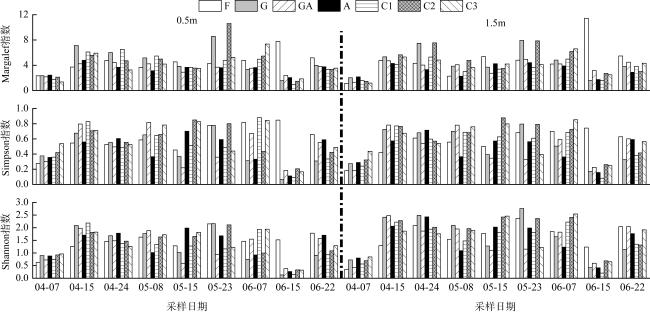
图5 南澳深澳湾海域不同类型调查区域浮游动物多样性指数的时空变化A: 鲍养殖区; C: 对照区; F: 鱼类养殖区; GA: 贝藻复合养殖区; G: 龙须菜栽培区 Fig. 5 Spatial-temporal changes in biodiversity indices of zooplankton in different areas of the Shen’ao Bay, Nan’ao Island. A: abalone mariculture area; C: natural sea area; F: fish mariculture area; GA: G. lemaneiformis-abalone integrated mariculture area; G: G. lemaneiformis cultivation area |
图6 南澳深澳湾海域不同类型调查区域中小型浮游动物体长(单位: mm)频度的时空变化A: 鲍养殖区; C: 对照区; F: 鱼类养殖区; GA: 贝藻复合养殖区; G: 龙须菜栽培区 Fig. 6 Spatial-temporal change in size-frequency distributions of zooplankton body length (units: mm) in different areas of the Shen’ao Bay, Nan’ao Island. A: Abalone mariculture area; C: natural sea area; F: fish mariculture area; GA: G. lemaneiformis-abalone integrated mariculture area; G: G. lemaneiformis cultivation area |
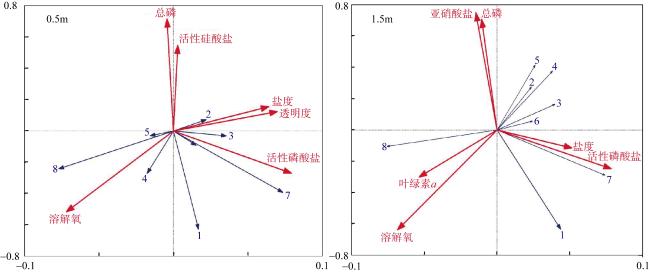
表2 南澳深澳湾海域不同类型调查区域0.5m层RDA排序前2轴与环境因子之间的相关系数Tab. 2 Correlation coefficients between the first 2 axes of RDA ranking and environmental factors at 0.5 m layers in different sea areas of the Shen’ao Bay, Nan’ao Island |
| 物种轴1 | 物种轴2 | 环境轴1 | 环境轴2 | |
|---|---|---|---|---|
| 物种轴1 | 1 | |||
| 物种轴2 | 0.0831 | 1 | ||
| 环境轴1 | 0.8934** | 0 | 1 | |
| 环境轴2 | 0 | 0.6816** | 0 | 1 |
| 透明度 | 0.6226** | 0.0847 | 0.697** | 0.1242 |
| 盐度 | 0.5711** | 0.1058 | 0.6392** | 0.1553 |
| 溶解氧 | -0.6419** | -0.3515** | -0.7185* | -0.5156** |
| 活性磷酸盐 | 0.7067** | -0.1833 | 0.7911** | -0.269* |
| 总磷 | -0.0381 | 0.4873** | -0.0427 | 0.715** |
| 活性硅酸盐 | 0.0262 | 0.3721** | 0.0293 | 0.5459** |
注: *P<0.05; **P<0.01。 表3 南澳深澳湾海域不同类型调查区域1.5m层RDA排序前2轴与环境因子之间的相关系数 |
Tab. 3 Correlation coefficients between the first 2 axes of RDA ranking and environmental factors at 1.5 m layers in different sea areas of the Shen’ao Bay, Nan’ao Island |
| 物种轴1 | 物种轴2 | 环境轴1 | 环境轴2 | |
|---|---|---|---|---|
| 物种轴1 | 1 | |||
| 物种轴2 | 0.0262 | 1 | ||
| 环境轴1 | 0.8919** | 0 | 1 | |
| 环境轴2 | 0 | 0.82** | 0 | 1 |
| 盐度 | 0.4578** | -0.0936 | 0.5133** | -0.1141 |
| 溶解氧 | -0.6092** | -0.5219** | -0.683** | -0.6365** |
| 叶绿素a | -0.4771** | -0.2454 | -0.5349** | -0.2992* |
| 亚硝酸盐 | -0.1297 | 0.6134** | -0.1454 | 0.748** |
| 活性磷酸盐 | 0.6989** | -0.2022 | 0.7836** | -0.2466 |
| 总磷 | -0.0945 | 0.5794** | -0.1059 | 0.7065** |
注: *P<0.05; **P<0.01。 |
图7 南澳深澳湾海域不同类型调查区域浮游动物常见种与环境因子的RDA分析图1. 小拟哲水蚤; 2. 小毛猛水蚤; 3. 尖额谐猛水蚤; 4. 长腹剑水蚤属; 5. 桡足类无节幼体; 6. 蔓足类六肢幼体; 7. 夜光虫; 8. 拟铃虫属 Fig. 7 RDA analysis between zooplankton species and environmental factors in different areas of the Shen’ao Bay, Nan’ao Island. 1. Paracalanus parvus; 2. Microsetella norvegica; 3. Euterpina acutifrons; 4. Oithona sp.; 5. nauplius larva (Copepod); 6. Cirripedia larva; 7. Noctiluca scintillans; 8. Tintinnopsis sp. |
| [1] |
陈清潮, 章淑珍, 1965. 黄海和东海的浮游桡足类Ⅰ. 哲水蚤目[J]. 海洋科学集刊, (7): 20-131.
|
| [2] |
陈清潮, 章淑珍, 1974a. 南海的浮游桡足类Ⅰ.[J]. 海洋科学集刊, (9): 101-116.
|
| [3] |
陈清潮, 沈嘉瑞, 1974b. 南海的浮游桡足类Ⅱ.[J]. 海洋科学集刊, (9): 126-137.
|
| [4] |
陈清潮, 章淑珍, 朱长寿, 1974c. 黄海和东海的浮游桡足类Ⅱ. 剑水蚤目和猛水蚤目[J]. 海洋科学集刊, (9): 27-76.
|
| [5] |
邓邦平, 杨宇峰, 2009. 大鹏澳养殖海域微表层与次表层水质及浮游动物群落特征的比较[J]. 暨南大学学报(自然科学与医学版), 30(1): 101-105.
|
| [6] |
刘萍, 宋洪军, 张学雷, 等, 2015. 桑沟湾浮游动物群落时空分布及养殖活动对其影响[J]. 海洋科学进展, 33(4): 501-511.
|
| [7] |
柳原, 柯志新, 李开枝, 等, 2024. 人类活动和沿岸流影响下的粤东近海浮游动物群落特征[J]. 热带海洋学报, 43(4): 98-111.
|
| [8] |
杜萍, 徐晓群, 徐旭丹, 等, 2017. 象山港三种不同养殖方式对浮游动物群落的影响[J]. 水产学报, 41(11): 1719-1733.
|
| [9] |
农业农村部渔业渔政管理局, 全国水产技术推广总站, 中国水产学会, 2022. 2022中国渔业统计年鉴[M]. 北京: 中国农业出版社: 22-23 (in Chinese).
|
| [10] |
任玉正, 柯志新, 谭烨辉, 等, 2020. 广东省南澳岛东部海域浮游动物群落结构及其影响因素[J]. 热带海洋学报, 39(2): 65-76.
|
| [11] |
苏家齐, 朱长波, 李俊伟, 等, 2019. 流沙湾浮游动物群落特征及与鱼贝养殖的关系[J]. 海洋渔业, 41(3): 278-293.
|
| [12] |
孙鲁峰, 李秀启, 徐兆礼, 2017. 东山湾浮游动物数量特征与养殖活动及水团关系分析[J]. 大连海洋大学学报, 32(4): 465-472.
|
| [13] |
徐淑敏, 齐占会, 史荣君, 等, 2019. 水产养殖对亚热带海湾氮磷营养盐时空分布的影响——以深澳湾为例[J]. 南方水产科学, 15(4): 29-38.
|
| [14] |
曾碧健, 岳晓彩, 黎祖福, 等, 2016. 生态浮床原位修复对海水养殖池塘浮游动物群落结构的影响[J]. 海洋与湖沼, 47(2): 354-359.
|
| [15] |
张武昌, 赵楠, 陶振铖, 等, 2010. 中国海浮游桡足类图谱[M]. 北京: 科学出版社.
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
/
| 〈 |
|
〉 |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}