
西沙群岛长棘海星(Acanthaster solaris)繁殖生物学的初步研究
|
刘界宏(1998—), 男, 辽宁省抚顺市人, 硕士研究生, 从事渔业生物学研究。email:liujiehong9898@163.com |
Editor: 林强
收稿日期: 2023-12-29
修回日期: 2024-02-22
网络出版日期: 2024-02-29
基金资助
国家重点研发计划(2022ZDYF3106303)
国家重点研发计划(2022YFD2401302)
国家自然科学基金项目(41876207)
国家自然科学基金项目(42376114)
海南省自然科学基金创新团队项目(322CXTD507)
国家自然科学基金地区项目(42166004)
Preliminary study on the reproductive biology of crown-of-thorns starfish (Acanthaster solaris) in the Xisha Islands
Editor: LIN Qiang
Received date: 2023-12-29
Revised date: 2024-02-22
Online published: 2024-02-29
Supported by
National Key Research and Development Program of China(2022ZDYF3106303)
National Key Research and Development Program of China(2022YFD2401302)
National Natural Science Foundation of China(41876207)
National Natural Science Foundation of China(42376114)
Hainan Provincial Natural Science Foundation Innovation Team Project(322CXTD507)
National Natural Science Foundation of China Regional Project(42166004)
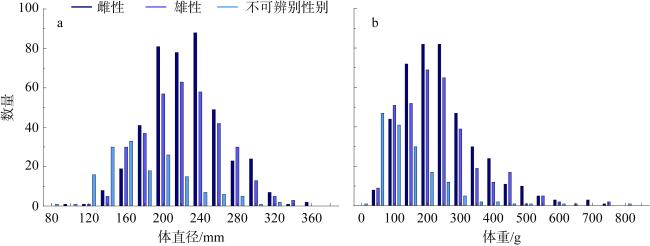
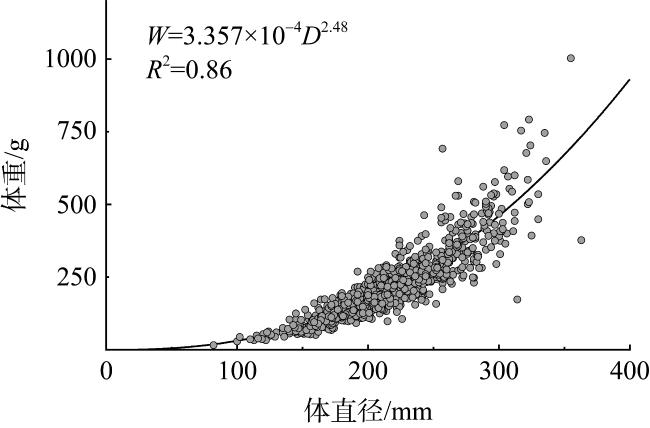
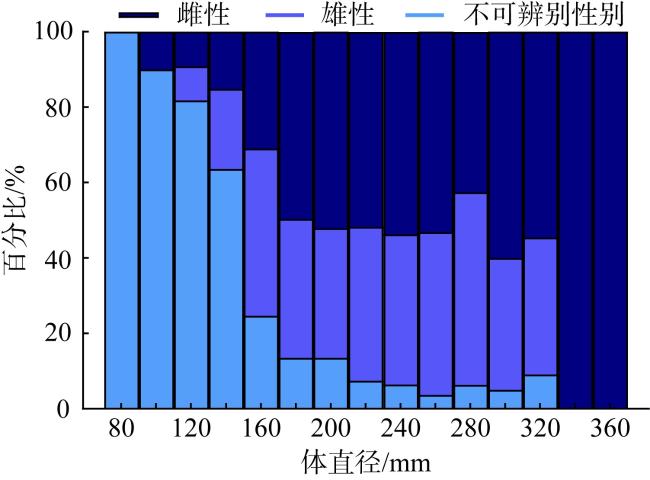
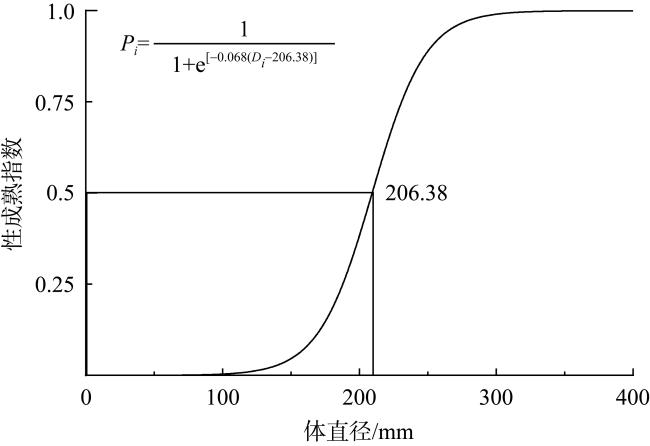
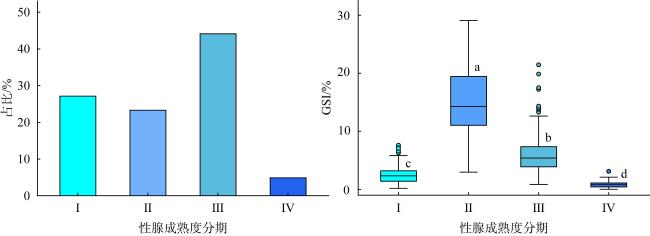
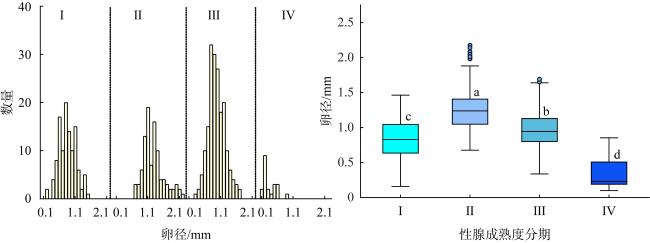
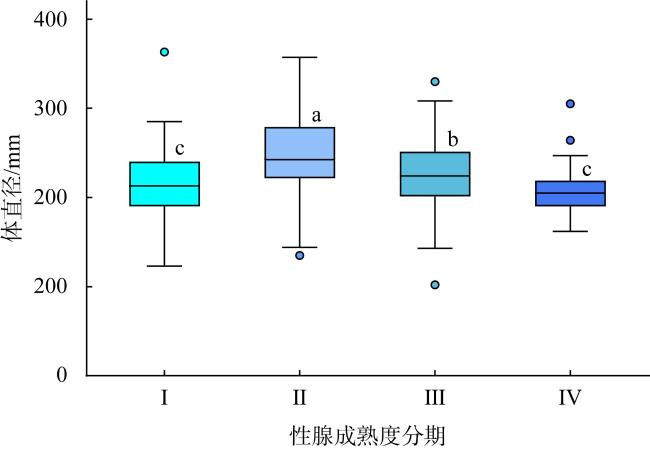
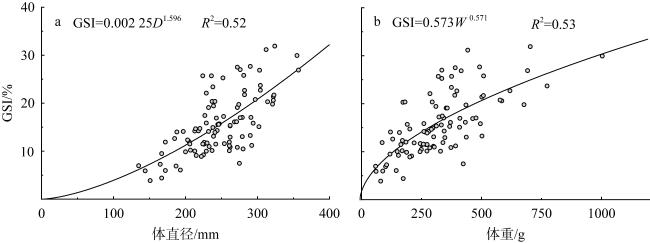
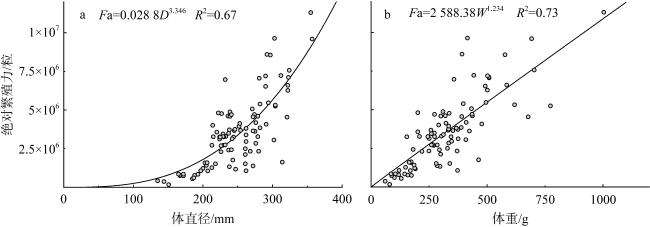
长棘海星(Acanthaster solaris)种群的暴发严重破坏了珊瑚礁生态系统, 对生物多样性造成了显著的影响。深入了解长棘海星的繁殖特征是预测其种群暴发和进行科学管理的关键。本研究于2022年5—7月在西沙群岛十个岛礁采集了长棘海星样品, 对其中928只开展了基础繁殖生物学研究。研究结果显示, 西沙群岛长棘海星的体直径(diameter, D)范围为82~363mm, 平均值为(217±45)mm, 体重(weight, W)范围为16.89~1 003.98g, 平均值为(224.69±123.6)g。体直径和体重呈幂函数关系, 关系式为W=3.357×10-4D2.48 (R2=0.86), 种群生长呈异速生长。雌雄个体性比为1.22:1。雌性海星的初次性成熟体直径为206.38mm。性腺成熟度分为四个时期, 其中性腺指数(gonadosomatic index, GSI)和卵径在Ⅱ期达到最高, 平均值分别为(15.68±6.35)%和(1.27±0.32)mm。绝对繁殖力范围为174 720~11 317 560粒, 平均值为(3 555 081±2 301 657)粒, 相对繁殖力范围为2 176.65~23 851.30粒·g-1, 平均值为(10 581.89±4 213.73)粒·g-1。绝对繁殖力与体直径和体重呈幂函数关系, 关系式分别为Fa=0.028 8D3.346 (R2=0.67, P<0.01)和Fa=2 588.38W1.234 (R2=0.73, P<0.01)。长棘海星的繁殖力与个体大小密切相关。本研究首次揭示了西沙群岛夏季长棘海星的繁殖特征, 将为了解长棘海星的种群动态和暴发机制提供科学依据, 对制定珊瑚礁生态系统有效的保护措施具有重要的意义。
刘界宏 , 王鹏程 , 王智豪 , 夏文彤 , 谢松光 , 宋一清 . 西沙群岛长棘海星(Acanthaster solaris)繁殖生物学的初步研究[J]. 热带海洋学报, 2024 , 43(6) : 160 -169 . DOI: 10.11978/2023203
The outbreak of the crown-of-thorns starfish (COTS, Acanthaster solaris) population has severely damaged coral reef ecosystems, leading to a significant impact on biodiversity. The knowledge of the reproductive characteristics of the COTS is crucial for predicting population outbreaks and scientific management strategies. In this study, the samples were collected from ten reefs in the Xisha Islands from May to July 2022, and a total of 928 individuals were conducted for the reproductive biology analysis. The results revealed that the body diameter (diameter, D) of the Xisha Islands ranges from 82 to 363 mm, the mean value was (217±45) mm, the body weight (weight, W) range was 16.89~1 003.98g, and the mean value was (224.69±123.6) g. The relationship between body diameter and weight followed a power function, expressed as W=3.357×10-4D2.48 (R2=0.86), indicating allometric growth in the population. The sex ratio of male to female individuals was 1.22:1. The initial sexual maturity body diameter for female COTS was 206.38mm. Gonadal maturity was classified into four stages, with the highest gonadosomatic index (GSI) and egg diameter occurring in Stage Ⅱ, averaging (15.68±6.35)%and (1.27±0.32) mm, respectively. The absolute fecundity range was 174 720~11 317 560 eggs, with mean of (3 555 081±2 301 657) eggs, relative fecundity range was 2 176.65~23 851.30 eggs·g-1, and the mean value was (10 581.89±4 213.73) eggs·g-1. The relationship between absolute fecundity and body diameter exhibited a power function (Fa=0.028 8D3.346, R2=0.67, P<0.01), and the relationship between relative fecundity and body weight exhibited a power function (Fa=2 588.38W1.234, R2=0.73, P<0.01). Reproductive capacity in COTS was closely related to individual size. This study, for the first time, reveals the reproductive characteristics of COTS in the Xisha Islands during the summer, providing a scientific basis for understanding the population dynamics and outbreak mechanisms of COTS. The results are significant for formulating effective conservation measures for coral reef ecosystems.
Key words: Xisha Islands; Acanthaster solaris; sex ratio; fecundity
表1 西沙群岛海域长棘海星样本采集信息Tab. 1 Sampling information for Acanthaster solaris in the Xisha Islands |
| 采集日期 | 岛礁 | 岛礁坐标 | 样本数量 |
|---|---|---|---|
| 2022年5月17日 | 全富岛 | 16°34′29.47″N, 111°40′24.05″E | 59 |
| 2022年5月20日 | 银屿 | 16°35′56.46″N, 111°42′7.27″E | 43 |
| 2022年5月21日 | 羚羊礁 | 16°28′59.41N, 111°35′29.47″E | 80 |
| 2022年5月21日 | 石屿 | 16°28′43.74″N, 111°35′45.82″E | 64 |
| 2022年5月21日 | 珊瑚岛 | 16°33′7.06″N, 111°37′32.35″E | 78 |
| 2022年5月24日 | 甘泉岛 | 16°30′40.44″N, 111°35′2.81″E | 72 |
| 2022年5月25日 | 金银岛 | 16°27′35.25″N, 111°31′20.03″E | 175 |
| 2022年7月22日 | 玉琢礁 | 16°19′24.37″N, 111°59′59.94″E | 66 |
| 2022年7月25日 | 浪花礁 | 16°3′26.50″N, 112°29′3.04″E | 115 |
| 2022年7月27日 | 东岛 | 16°40′5.85″N, 112°44′36.34″E | 176 |
图4 2022年5月至7月西沙群岛海域长棘海星的性别比例Fig. 4 Gender ratio of Acanthaster solaris in the Xisha Islands from May to July 2022 |
图6 2022年5月至7月西沙群岛海域长棘海星雌性性腺成熟度占比与性腺指数Fig. 6 Percentage of female gonads and gonadosomatic index (GSI) at different development stages of Acanthaster solaris in the Xisha Islands from May to July 2022 |
图7 2022年5月至7月西沙群岛海域长棘海星雌性性腺各成熟度时期卵径大小频数分布与卵径大小Fig. 7 Oocyte size-frequency distribution and oocyte size at different development stages of Acanthaster solaris in the Xisha Islands from May to July 2022 |
| [1] |
蔡健, 梁曼颖, 王健娇, 等, 2021. 长棘海星中含氮类化学成分研究[J]. 中国海洋药物, 40(4): 45-49.
|
| [2] |
段友健, 2015. 拉萨裸裂尻鱼个体生物学和种群动态研究[D]. 武汉: 华中农业大学.
|
| [3] |
李渊, 胡战彪, 谢世君, 等, 2024. 南海长棘海星物种有效性探讨[J]. 应用海洋学学报, 43(2): 201-207.
|
| [4] |
李元超, 吴钟解, 梁计林, 等, 2019a. 近15年西沙群岛长棘海星暴发周期及暴发原因分析[J]. 科学通报, 64(33): 3478-3484.
|
| [5] |
李元超, 梁计林, 吴钟解, 等, 2019b. 长棘海星的暴发及其防治[J]. 海洋开发与管理, 36(8): 9-12.
|
| [6] |
刘晨斌, 徐革锋, 黄天晴, 等, 2019. 鱼类性腺发育研究进展[J]. 水产学杂志, 32(1): 46-54.
|
| [7] |
王锦溪, 邓裕坚, 范瀛方, 等, 2023. 北部湾多齿蛇鲻繁殖生物学特征[J]. 上海海洋大学学报, 32(4): 794-805.
|
| [8] |
严林, 田静, 彭祖想, 等, 鸭绿江丹东段扁吻鮈的个体生殖力及初次性成熟体长与年龄[J]. 大连海洋大学学报, 2023, 38(1): 112-119.
|
| [9] |
张颖, 杨栎潼, 刘冰, 等, 2022. 长棘海星(Acanthaster planci)幼体特异性PCR检测技术与应用[J]. 热带海洋学报, 41(6): 125-131.
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}