
粤东上升流区不同水层可培养细菌多样性
|
张敬(1983—), 女, 工程师, 硕士。海洋环境微生物学。email: zhangjing@scsio.ac.cn |
Editor: 林强
收稿日期: 2023-12-05
修回日期: 2024-03-29
网络出版日期: 2024-04-03
基金资助
广东省科技计划项目(2021B1212050023)
广东省“珠江人才计划”本土创新科研团队项目(2019BT02Y262)
Diversity of culturable bacteria in different water layers in the upwelling area of eastern Guangdong
Editor: LIN Qiang
Received date: 2023-12-05
Revised date: 2024-03-29
Online published: 2024-04-03
Supported by
Science and Technology Planning Project of Guangdong Province, China(2021B1212050023)
Local Innovative and Research Teams Project of Guangdong Pearl River Talents Program(2019BT02Y262)
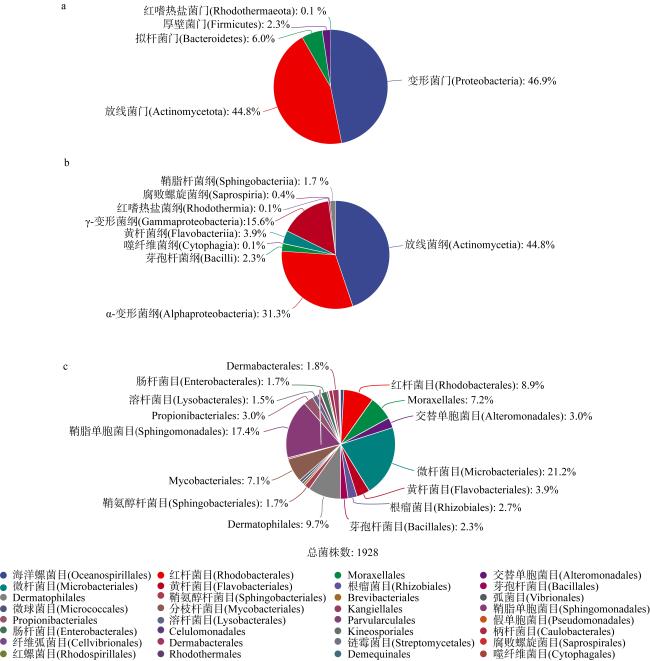
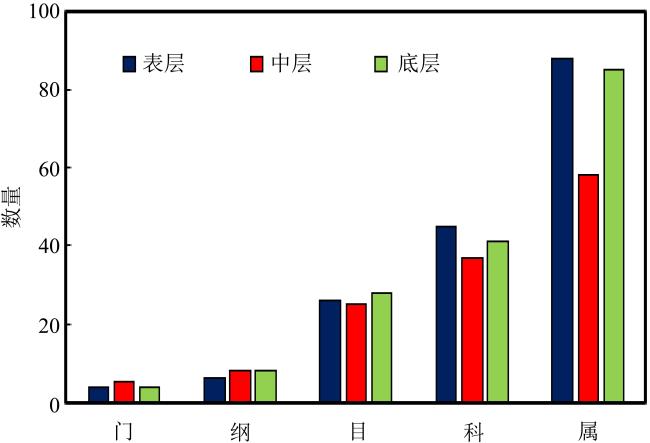
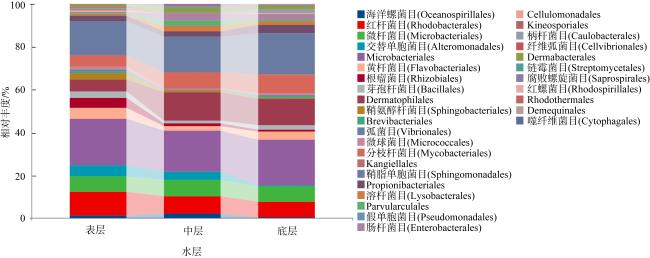
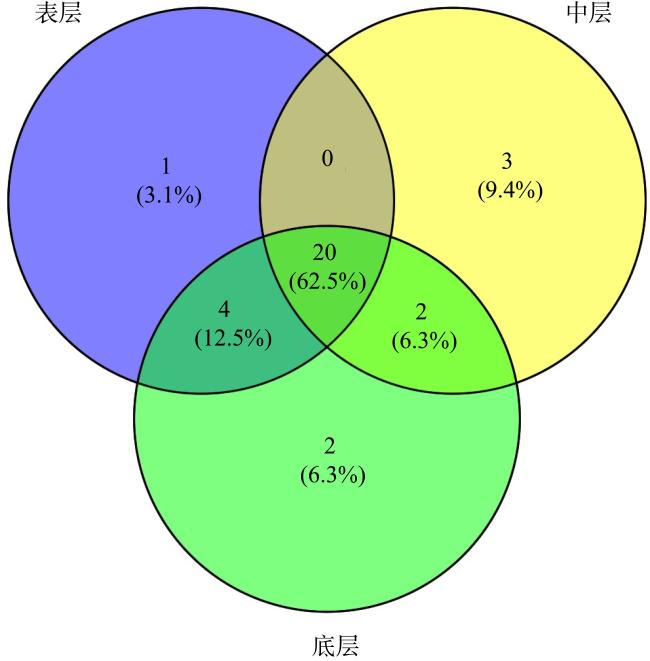
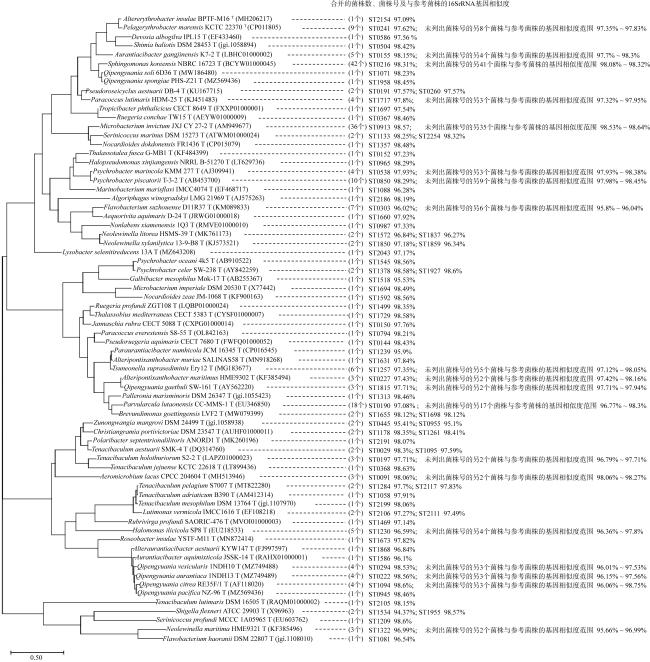
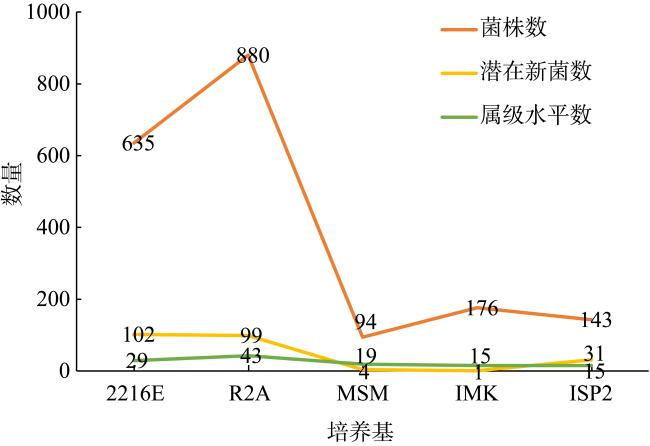
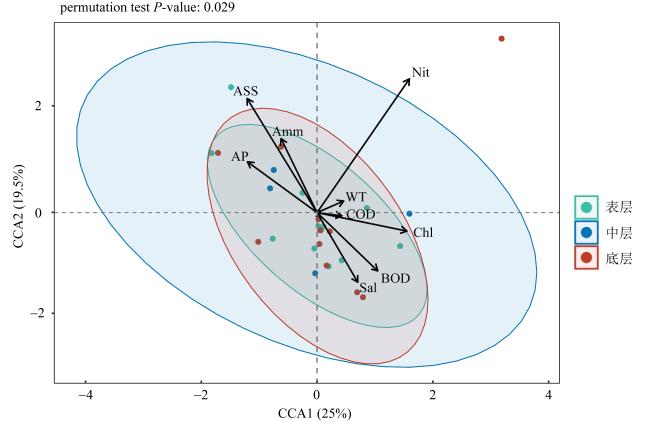
上升流通过增强扩散和独特的环境驱动着近海生态系统中的物质循环和能量流动, 直接影响环境中生物的生长、繁殖和分布, 蕴藏了大量尚未培养和难培养的微生物和新基因资源。本研究从粤东上升流区表层、中层和底层共采集27份海水样品, 探究细菌多样性和分布模式及其与深度的相关性。采用纯培养方法共分离获得1928株细菌, 通过16S rRNA基因序列分析, 结果显示这些细菌分布在5门、9纲、32目、54科、121属中的262种。其中, 变形菌门(Proteobacteria)为优势门, 占46.9%; 放线菌纲(Actinomycetia)为优势纲, 占44.8%; Microbacteriales为优势目级类群, 占21.2%。总体来说, 中层海水分离得到的菌株多样性明显低于表层和底层海水环境, 但三个层级环境分离获得的优势种群基本相同, 最优类群均为Microbacteriales目。从不同水层海水中分离得到的潜在新菌数排序为: 底层>表层>中层。从不同培养基分离得到的潜在新菌率排序为2216E培养基>R2A培养基>ISP2培养基>MSM培养基>IMK培养基。通过细菌多样性与环境因子的典范对应分析 (canonical correspondence analysis, CCA)表明, 能显著影响细菌多样性变化的环境因子是叶绿素含量(chlorophyll content, Chl)、活性硅酸盐(active silicate salts, ASS)、活性磷酸盐 (active phosphate, AP)、氨及部分氨基酸(ammonia, Amm)和生化需氧量(biochemical oxygen demand, BOD)。本研究系统调查了粤东上升流区可培养海洋细菌多样性及其分布情况, 其结果为近海上升流区不同水层可培养细菌多样性研究积累了关键的资料和数据, 也为近海典型上升流环境生物物种和基因资源的发掘和利用提供了重要参考。
张敬 , 凌娟 , 石松标 , 韦章良 , 范利可 . 粤东上升流区不同水层可培养细菌多样性[J]. 热带海洋学报, 2024 , 43(6) : 80 -91 . DOI: 10.11978/2023186
Upwelling promotes material cycling and energy flow in offshore ecosystems through enhanced diffusion and unique environmental drivers, which directly affects the growth, reproduction, and distribution of organisms in the environment. In this environment there are a large number of uncultivated or difficult to cultivate microorganisms and new gene resources, which need to be cultured and explored in deeply. In this study, 27 seawater samples were collected from the surface, middle and bottom layers of the upwelling area in eastern Guangdong to explore the diversity and distribution patterns of bacteria and their correlation with the depth. A total of 1928 bacterial strains were isolated by pure culture method, and identified by 16S rRNA gene sequences. Results showed that these bacteria were distributed in 5 phyla, 9 classes, 32 orders, 54 families, 121 genera and 262 species. Among them, Proteobacteria was the dominant phylum, accounting for 46.9%. Actinomycetia was the dominant class, accounting for 44.8%. Microbacteriales was the dominant order, accounting for 21.2%. In general, the diversity of species from the mid-level seawater was significantly lower than that from the surface and bottom seawater environments, but the dominant populations obtained from the three levels were basically the same, and the most optimal group was Microbacteriales. The number of potential new bacteria isolated from different water layers was ranked as bottom layer> surface layer> middle layer. The rate of potential new bacteria isolated from different media was ranked as follows of 2216E medium> R2A medium> ISP2 medium> MSM medium> IMK medium. CCA (canonical correspondence analysis) showed that the environmental factors that significantly affected the changes of bacterial diversity were Chl (chlorophyll content), ASS (active silicate salts), AP (active phosphate), Amm (ammonia) and BOD (biochemical oxygen demand). In this study, we systematically investigated the diversity and distribution of culturable marine bacterial in the upwelling area of eastern Guangdong. These results showed the key microbial information and data for the further study on culturabble bacterial diversity in the offshore upwelling area, and also provided reference for the exploration and utilization of species and gene resources in the typical ocean environment.
表1 样品采集的站位Tab. 1 Sampling sites |
| 调查站点 | 取样纬度 | 取样经度 | 测站水深/m | 表层 | 中层 | 底层 |
|---|---|---|---|---|---|---|
| S01 | 23˚33'17"N | 117˚03'27"E | 11 | √ | √ | |
| S02 | 23˚31'68"N | 117˚03'55"E | 8 | √ | √ | |
| S03 | 23˚28'40"N | 116˚55'83"E | 5 | √ | ||
| S04 | 23˚28'95"N | 116˚59'46"E | 14 | √ | √ | |
| S05 | 23˚29'01"N | 117˚11'91"E | 15 | √ | √ | |
| S06 | 23˚24'49"N | 117˚09'32"E | 29 | √ | √ | √ |
| S07 | 23˚23'26"N | 117˚22'15"E | 40 | √ | √ | √ |
| S08 | 23˚15'31"N | 117˚18'35"E | 39 | √ | √ | √ |
| S09 | 23˚18'39"N | 117˚04'55"E | 19 | √ | √ | |
| S10 | 23˚21'41"N | 116˚50'58"E | 8.5 | √ | √ | |
| S11 | 23˚11'27"N | 116˚53'36"E | 27 | √ | √ | √ |
| S12 | 23˚16'36"N | 116˚47'94"E | 9.2 | √ | √ |
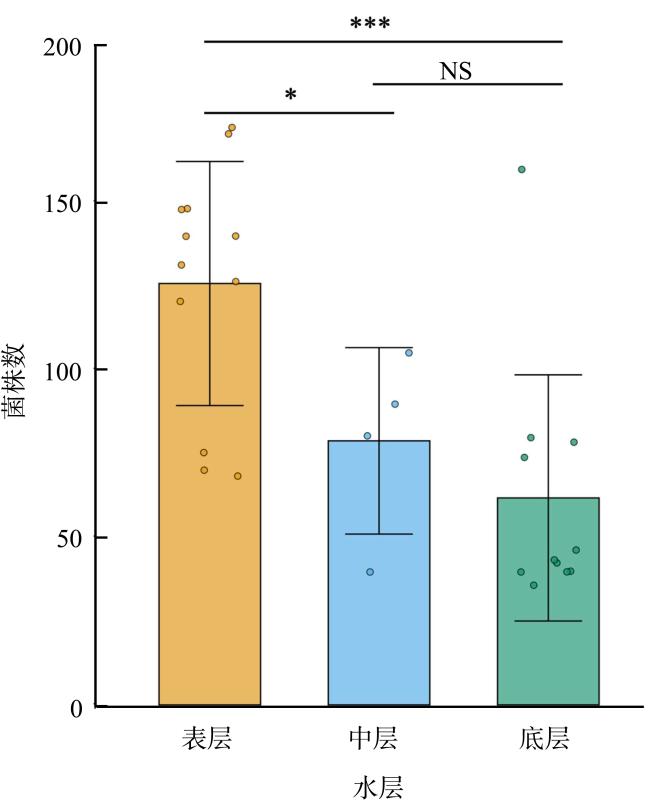
图1 不同水层菌落数比较圆点为每个水层原样海水稀释至10-2后100μL菌悬液培养出的菌落数, 配对样本t检验, ***P<0.001; **P<0.01; *P<0.05; NS, P> 0.05 Fig. 1 Comparison of the number of colonies in different water layers. The number of colonies cultured from 100μL bacterial suspension after dilution of the original seawater to 10-2, Paired-Samples t test, ***P<0.001; **P<0.01; *P<0.05; NS, P>0.05 |
表2 所有海水样品环境参数Tab. 2 Environmental parameters of all seawater samples |
| 站点 | 采样层 | 环境参数 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DO/ (mg·L-1) | Chl/ (μg·L-1) | WT/℃ | Sal/‰ | SS/ (mg·L-1) | Tur/ TU | pH | Tra / m | ASS /(mg·L-1) | AP/ (mg·L-1) | Sub/ (mg·L-1) | Nit/ (mg·L-1) | Amm/ (mg·L-1) | COD/ (mg·L-1) | BOD/ (mg·L-1) | ||
| S01 | 表 | 6.083 | 0.569 | 23.8 | 30.4 | 5.60 | 7.57 | 7.91 | 1.7 | 1.07 | 0.049 | 0.024 | 0.043 | 0.138 | 1.048 | 0.494 |
| 底 | 6.225 | 0.498 | 23.6 | 30.9 | 11.80 | 13.33 | / | / | 0.54 | 0.030 | 0.025 | 0.034 | 0.125 | 0.944 | 0.497 | |
| S02 | 表 | 6.744 | 1.802 | 23.4 | 31.2 | 10.55 | 4.56 | 7.99 | 2.1 | 0.39 | 0.031 | 0.013 | 0.054 | 0.052 | 0.942 | 0.250 |
| 底 | 6.646 | 1.027 | 23.2 | 30.9 | 10.55 | 3.80 | / | / | 0.56 | 0.043 | 0.013 | 0.039 | 0.056 | 1.142 | 0.184 | |
| S03 | 表 | 6.992 | 0.774 | 23.1 | 31.2 | 10.70 | 5.81 | 8.02 | 1.2 | 0.63 | 0.008 | 0.027 | 0.003 | 0.027 | 0.852 | 0.644 |
| S04 | 表 | 6.604 | 0.322 | 23.0 | 31.1 | 12.15 | 8.71 | 8.00 | 1.3 | 0.82 | 0.017 | 0.019 | 0.014 | 0.057 | 0.978 | 0.115 |
| 底 | 6.706 | 0.409 | 23.0 | 30.8 | 13.60 | 12.71 | / | / | 0.65 | 0.008 | 0.021 | 0.037 | 0.051 | 0.703 | 0.395 | |
| S05 | 表 | 7.161 | 0.340 | 22.6 | 33.4 | 4.60 | 1.96 | 8.13 | 2.9 | 0.35 | 0.007 | 0.003 | 0.003 | 0.008 | 1.003 | 0.344 |
| 底 | 6.898 | 0.607 | 22.6 | 33.5 | 5.53 | 4.57 | / | / | 0.37 | 0.008 | 0.003 | 0.003 | 0.030 | 1.098 | 0.151 | |
| S06 | 表 | 6.722 | 0.559 | 22.7 | 33.3 | 4.20 | 2.20 | 8.14 | 2 | 0.44 | 0.009 | 0.007 | 0.003 | 0.055 | 0.796 | 0.312 |
| 中 | 6.630 | 0.605 | 22.7 | 33.3 | 8.65 | 7.32 | / | / | 0.40 | 0.010 | 0.007 | 0.003 | 0.043 | 0.828 | 0.268 | |
| 底 | 6.692 | 0.658 | 22.7 | 33.3 | 9.00 | 5.86 | / | / | 0.35 | 0.008 | 0.006 | 0.003 | 0.023 | 0.849 | 0.302 | |
| S07 | 表 | 6.761 | 0.153 | 23.0 | 34.0 | 0.60 | 3.07 | 8.11 | 10.2 | 0.18 | 0.007 | 0.003 | 0.003 | 0.022 | 0.479 | 0.013 |
| 中 | 7.023 | 0.501 | 23.0 | 34.0 | 0.60 | 1.60 | / | / | 0.16 | 0.006 | 0.003 | 0.003 | 0.006 | 0.950 | 0.096 | |
| 底 | 6.813 | 0.600 | 23.0 | 34.0 | 0.95 | 1.13 | / | / | 0.12 | 0.008 | 0.003 | 0.003 | 0.047 | 0.496 | 0.029 | |
| S08 | 表 | 6.926 | 0.499 | 23.3 | 34.1 | 0.95 | 3.35 | 8.15 | 11.2 | 0.08 | 0.008 | 0.003 | 0.003 | 0.003 | 0.772 | 0.010 |
| 中 | 6.893 | 0.414 | 23.3 | 34.1 | 1.15 | 2.24 | / | / | 0.11 | 0.006 | 0.003 | 0.003 | 0.007 | 0.584 | 0.108 | |
| 底 | 6.758 | 0.401 | 23.3 | 34.1 | 1.10 | 0.83 | / | / | 0.19 | 0.008 | 0.003 | 0.003 | 0.010 | 0.772 | 0.027 | |
| S09 | 表 | 8.898 | 5.786 | 23.3 | 32.5 | 2.45 | 2.84 | 8.28 | 3.4 | 0.23 | 0.006 | 0.003 | 0.003 | 0.007 | 1.297 | 0.829 |
| 底 | 7.475 | 3.084 | 22.8 | 33.5 | 3.70 | 3.48 | / | / | 0.24 | 0.007 | 0.003 | 0.003 | 0.003 | 1.149 | 0.383 | |
| S10 | 表 | 7.946 | 5.661 | 23.9 | 28.3 | 5.40 | 4.44 | 8.17 | 3.1 | 0.26 | 0.007 | 0.018 | 0.063 | 0.028 | 1.057 | 1.384 |
| 底 | 6.729 | 1.614 | 22.8 | 33.1 | 14.10 | 15.05 | / | / | 0.49 | 0.007 | 0.016 | 0.131 | 0.046 | 0.801 | 0.693 | |
| S11 | 表 | 7.557 | 0.466 | 23.5 | 33.4 | 4.40 | 0.82 | 8.14 | 5.2 | 0.23 | 0.005 | 0.003 | 0.003 | 0.007 | 0.853 | 0.835 |
| 中 | 7.129 | 0.555 | 23.1 | 33.8 | 1.25 | 1.13 | / | / | 0.24 | 0.006 | 0.003 | 0.003 | 0.012 | 1.331 | 0.244 | |
| 底 | 6.911 | 0.766 | 23.2 | 33.5 | 2.95 | 4.99 | / | / | 0.23 | 0.006 | 0.003 | 0.003 | 0.008 | 0.752 | 0.113 | |
| S12 | 表 | 7.429 | 1.206 | 23.8 | 27.5 | 3.40 | 4.82 | 8.08 | 2.7 | 0.67 | 0.008 | 0.024 | 0.003 | 0.041 | 1.429 | 1.333 |
| 底 | 7.412 | 0.914 | 22.9 | 33.3 | 6.45 | 7.69 | / | / | 0.71 | 0.008 | 0.015 | 0.008 | 0.063 | 0.916 | 1.196 | |
注: pH、Tra为根据站位监测站收集的固定数据,未分层 |
图8 可培养细菌多样性与叶绿素含量(Chl)、水温(WT)、盐度(Sal)、活性硅酸盐(ASS)、活性磷酸盐(AP)、硝酸盐(Nit)、氨及部分氨基酸(Amm)、化学需氧量(COD)和生化需氧量(BOD)的典范对应分析Fig. 8 Canonical correspondence analysis(CCA) between diversity of culturable bacteria and chlorophyll content (Chl), water temperature (WT), salinity(Sal), active silicate salts (ASS), active phosphate (AP), nitrate (Nit), ammonia and some amino acids (Amm), chemical oxygen demand (COD) and biochemical oxygen demand (BOD) |
表3 环境因子对可培养细菌多样性的ANOSIM检验结果(P-值)Tab. 3 ANOSIM test results (P-value) of environmental factors on changes of diversity of cultured bacteria |
| 环境因子 | P |
|---|---|
| 叶绿素含量(Chl) | 0.026 |
| 水温(WT) | 0.243 |
| 盐度(Sal) | 0.068 |
| 活性硅酸盐(ASS) | 0.011 |
| 活性磷酸盐(AP) | 0.002 |
| 硝酸盐(Nit) | 0.866 |
| 氨及部分氨基酸(Amm) | 0.001 |
| 化学需氧量(COD) | 0.150 |
| 生化需氧量(BOD) | 0.001 |
| [1] |
洪启明, 李立, 1991. 粤东陆架区夏季的上升流[J]. 台湾海峡, 10(3): 271-277.
|
| [2] |
乐凤凤, 蔡昱明, 孙军, 等, 2015. 2009年冬、夏季南海北部超微型浮游生物的分布特征及其环境相关分析[J]. 海洋学报, 37(12): 41-55.
|
| [3] |
李存, 崔林青, 杨红强, 等, 2022. 三份南海岛礁珊瑚砂样品中可培养细菌多样性[J]. 热带海洋学报, 41(2): 149-158.
|
| [4] |
李涵, 樊成奇, 陈莎, 等, 2022. 南海北部表层海水微生物多样性及部分真菌活性物质研究[J]. 海洋渔业, 44(4): 446-458.
|
| [5] |
李立, 1990. 珠江口以西陆架夏季上升流的研究[J]. 台湾海峡, 9(4): 338-346.
|
| [6] |
毛梦婷, 张瑾, 文姣, 等, 2023. 南极乔治王岛冰锥洞微生物培养探索[J]. 微生物学报, 63(6): 2066-2077.
|
| [7] |
商丽, 史晓翀, 王晓宇, 等, 2017. 北欧海海水可培养细菌多样性[J]. 微生物学报, 57(9): 1323-1331.
|
| [8] |
孙创, 王金燕, 张钰琳, 等, 2021. 利用改良培养基探究西太平洋海水可培养细菌多样性[J]. 微生物学报, 61(4): 845-861.
|
| [9] |
续子杰, 肖鹏, 杨义刚, 等, 2021. 淡水浮游细菌群落采集、过滤与保存[J]. 微生物组实验手册, Bio-101: e2003736.
|
| [10] |
薛璐, 向冬芳, 鲜博, 等, 2023. 保安湖水体细菌群落结构时空变化特征及驱动因子[J]. 水生生物学报, 47(3): 450-461.
|
| [11] |
张坤, 闫畅, 田新朋, 2023. 微生物单细胞分离方法研究进展[J]. 生物技术通报, 39(9): 1-11.
|
| [12] |
张晓华, 2007. 海洋微生物学[M]. 青岛: 中国海洋大学出版社.
|
| [13] |
周天旭, 罗文磊, 笪俊, 等, 2022. 抚仙湖垂向分层期间水体细菌群落结构组成及多样性的空间分布[J]. 湖泊科学, 34(5): 1642-1655.
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}