
珊瑚来源潜在降解对甲氧基肉桂酸辛酯的细菌物种多样性
|
黄沁愉(1999—), 女, 广东省广州市人, 硕士研究生, 从事海洋珊瑚微生物相关研究。email: hyhqy172@sina.com |
Copy editor: 孙翠慈
收稿日期: 2024-03-18
修回日期: 2024-04-12
网络出版日期: 2024-04-29
基金资助
国家自然科学基金项目(U23A2036)
Microbial diversity of potential EHMC-degrading bacteria from coral
Copy editor: SUN Cuici
Received date: 2024-03-18
Revised date: 2024-04-12
Online published: 2024-04-29
Supported by
National Natural Science Foundation of China(U23A2036)
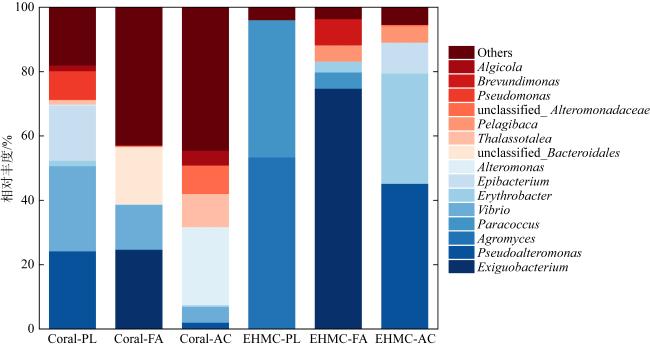
对甲氧基肉桂酸辛酯(2-ethylhexyl-4-methoxycinnamic acid, EHMC)是国内防晒产品中最常见的有机紫外吸收剂成分, 随人类活动汇入海洋并在珊瑚组织内积累, 加速珊瑚白化。微生物降解法是去除珊瑚EHMC污染的有效手段之一, 然而目前珊瑚的EHMC降解细菌资源及EHMC对珊瑚微生物群落的影响仍知之甚少。本文选择三亚鹿回头近岸海域珊瑚为研究对象, 开展EHMC降解菌多样性研究。首先, 以EHMC为唯一碳源对珊瑚组织样品进行多次富集, 获得EHMC的耐受菌群, 利用高通量测序技术, 分析富集菌群组成和多样性; 同时通过细菌的分离鉴定和功能验证, 筛选出潜在的EHMC降解菌株。16S rRNA基因高通量测序结果显示, 在EHMC胁迫下, 滨珊瑚(Porites lutea)、角蜂巢珊瑚(Favosites sp.)和鹿角珊瑚(Acropora sp.)三个样品的细菌群落多样性下降。从属水平上看, 壤霉菌属(Agromyces)、副球菌属(Paracoccus)、Pelagibaca属和赤杆菌属(Erythrobacter)等细菌在原始珊瑚中占比极低, 经EHMC多次富集后成为优势菌属。通过纯化培养, EHMC富集样品共分离出60株可培养细菌。16S rRNA基因鉴定结果显示, 60株细菌分别归属于3个门、3个纲、6个目、8个科、13个属和17个种。可培细菌的优势菌属主要为芽孢杆菌属(Bacillus), 其次是红球菌属(Rhodococcus), 副球菌属, 微小杆菌属(Exiguobacterium)和尖球菌属(Acuticoccus)。经对单个纯培养菌株的活性筛选, 最终筛选出8株潜在EHMC降解菌株, 分别归属于芽孢杆菌属、短波单胞菌属(Brevundimonas)、副球菌属、尖球菌属、Phycicoccus属、红球菌属和分枝杆菌属(Mycolicibacterium)。前人研究表明, 红球菌属、芽孢杆菌属和副球菌属等细菌与芳香族化合物的生物降解相关。本研究分析了EHMC污染对珊瑚细菌群落的影响, 并分离出潜在的EHMC降解细菌, 为珊瑚抵御有机紫外吸收剂污染提供有用信息, 也为后续构建珊瑚益生微生物组减轻珊瑚礁有机污染积累了宝贵的菌种资源。
黄沁愉 , 吕丽娜 , 李洁 , 琚慧敏 , 苏宏飞 . 珊瑚来源潜在降解对甲氧基肉桂酸辛酯的细菌物种多样性[J]. 热带海洋学报, 2025 , 44(1) : 133 -145 . DOI: 10.11978/2024059
2-ethylhexyl-4-methoxycinnamic acid (EHMC) is an organic UV filter that is widely used in domestic sunscreen products. It is released into the ocean via human activities and accumulated in coral tissues, hastening coral bleaching. Biodegradation is an effective method for removing EHMC from coral, but little is known about its biodegradation in coral and its impact on coral microorganisms. To study the diversity of EHMC biodegradation in coral, we collected coral samples from Luhuitou, Sanya. These samples were enriched multiple times with EHMC as the sole carbon source to obtain EHMC-tolerance bacteria. High-throughput sequencing technology was used to analyze the composition and diversity of bacteria communities from enrichment samples. Besides, potential EHMC-degrading strains were isolated and identified from the enrichment through activity tests. High-throughput 16S rRNA gene sequencing revealed that the microbial community diversity in coral samples of Porites lutea, Favosites sp. and Acropora sp. was reduced under high concentration of EHMC. In contrast to their low abundance in the original coral samples, Agromyces, Paracoccus, Pelagibaca and Erythrobacter were the dominant genera after multiple enrichment with EHMC. Furthermore, a total of 60 bacterial strains were isolated from the EHMC-enriched samples. Based on 16S rRNA gene identification, these strains belonged to three phyla, three classes, six orders, eight families, thirteen genera and seventeen species. The main dominant genera among the cultivable bacteria were Bacillus, followed by Rhodococcus, Paracoccus, Exiguobacterium and Acuticoccus. Moreover, eight potential EHMC-degrading strains were selected through activity tests, belonging to Bacillus, Brevundimonas, Paracoccus, Acuticoccus, Phycicoccus, Rhodococcus and Mycolicibacterium, respectively. Previous studies have shown that Rhodococcus, Bacillus and Paracoccus were correlated with the biodegradation of aromatic compounds. This study illustrates the impact of EHMC on coral microbial communities and isolates potential EHMC-degrading bacteria, providing valuable information for coral resilience against organic UV filters. Additionally, it accumulates valuable bacterial resources for eliminating organic contaminants in coral by the subsequent construction of coral probiotic consortium.
表1 三亚鹿回头海域采集的珊瑚物种信息Tab.1 The information of coral species collected from Luhuitou, Sanya |
| 珊瑚属名 | 珊瑚种名 | 拉丁名 |
|---|---|---|
| 蔷薇珊瑚属 | 截顶蔷薇珊瑚 | Montipora truncata |
| 蔷薇珊瑚 | Montipora sp. | |
| 盘星珊瑚属 | 盘星珊瑚 | Dipsastraea sp. |
| 牡丹珊瑚属 | 厚板牡丹珊瑚 | Pavona duerdeni |
| 十字牡丹珊瑚 | Pavona decussata | |
| 鹿角珊瑚科属 | 巨锥鹿角珊瑚 | Acropora monticulosa |
| 多孔鹿角珊瑚 | Acropora millepora | |
| 风信子鹿角珊瑚 | Acropora hyacinthus | |
| 鹿角珊瑚 | Acropora sp. | |
| 盔形珊瑚属 | 从生盔珊瑚 | Galaxea fascicularis |
| 角孔珊瑚属 | 角孔珊瑚 | Goniopora sp. |
| 角蜂巢珊瑚属 | 角蜂巢珊瑚 | Favosites sp. |
| 滨珊瑚属 | 澄黄滨珊瑚 | Porites lutea |
| 杯形珊瑚属 | 鹿角杯形珊瑚 | Pocillopora damicornis |
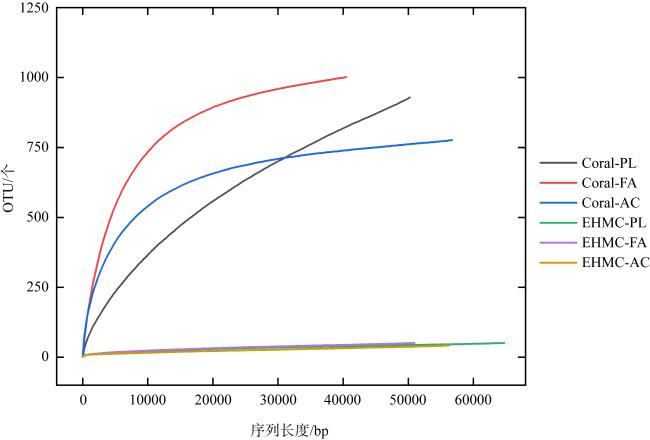
表2 原始珊瑚样品和EHMC富集样品的微生物群落多样性指数Tab. 2 Diversity indexes of microbial communities isolated from original coral and EHMC-enrichment samples |
| 珊瑚物种 | 样品编号 | ACE指数 | Chao1指数 | Simpson 指数 | Shannon 指数 |
|---|---|---|---|---|---|
| 滨珊瑚 | Coral-PL | 1888 | 1423 | 0.84 | 3.88 |
| EHMC-PL | 75 | 95 | 0.53 | 1.34 | |
| 角蜂巢珊瑚 | Coral-FA | 1040 | 1052 | 0.90 | 5.45 |
| EHMC-FA | 84 | 69 | 0.43 | 1.53 | |
| 鹿角珊瑚 | Coral-AC | 808 | 837 | 0.93 | 5.92 |
| EHMC-AC | 82 | 81 | 0.67 | 1.98 |
表3 原始珊瑚样品和EHMC富集样品中不同水平上的细菌种类数目Tab. 3 The numbers of bacteria species at different levels in original coral and EHMC-enrichment samples |
| 珊瑚物种 | 样品编号 | 门数目 | 纲数目 | 目数目 | 科数目 | 属数目 |
|---|---|---|---|---|---|---|
| 滨珊瑚 | Coral-PL | 26 | 48 | 118 | 193 | 335 |
| EHMC-PL | 5 | 7 | 21 | 33 | 43 | |
| 角蜂巢珊瑚 | Coral-FA | 31 | 64 | 154 | 250 | 394 |
| EHMC-FA | 6 | 8 | 20 | 31 | 40 | |
| 鹿角珊瑚 | Coral-AC | 19 | 38 | 99 | 174 | 346 |
| EHMC-AC | 4 | 6 | 16 | 25 | 37 |
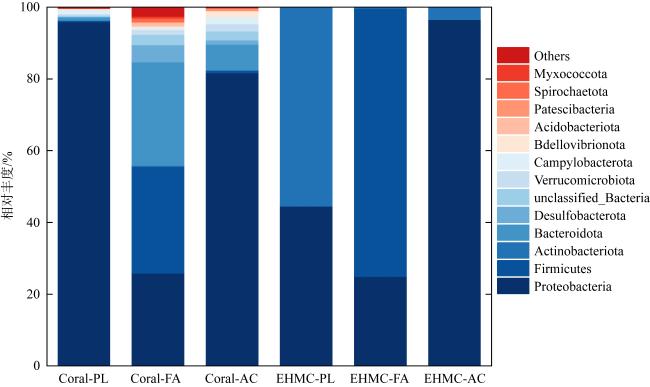
图2 原始珊瑚/EHMC富集样品在门水平上的细菌群落分布图中仅列出相对丰度> 0.1%的菌属类群; Others为相对丰度< 0.1%的菌属类群; 前缀中Coral-代表原始珊瑚样品, EHMC-代表富集样品; 后缀中PL代表滨珊瑚, FA代表角蜂巢珊瑚, AC代表鹿角珊瑚 Fig. 2 Distribution of bacterial community in original coral samples and EHMC-enrichment samples based on phylum level (relative abundance > 0.1%) |
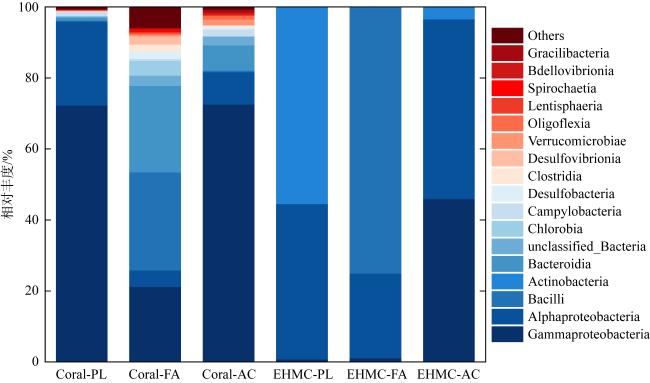
图3 原始珊瑚/EHMC富集样品在纲水平上的细菌群落分布图中仅列出相对丰度> 0.1%的菌属类群; Others为相对丰度< 0.1%的菌属类群; 前缀中Coral-代表原始珊瑚样品, EHMC-代表富集样品; 后缀中PL代表滨珊瑚, FA代表角蜂巢珊瑚, AC代表鹿角珊瑚 Fig. 3 Distribution of bacterial community in original coral samples and EHMC-enrichment samples based on class level (relative abundance > 0.1%) |
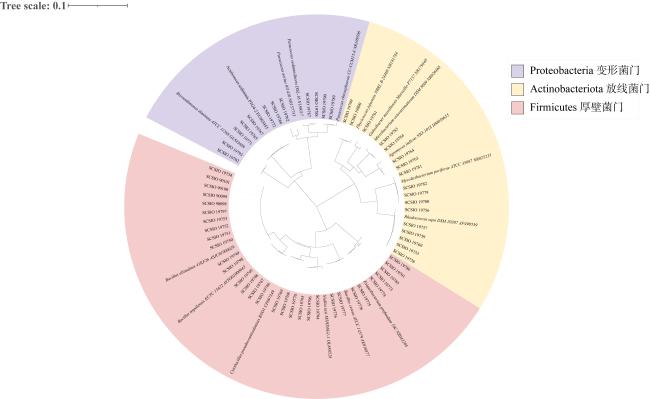
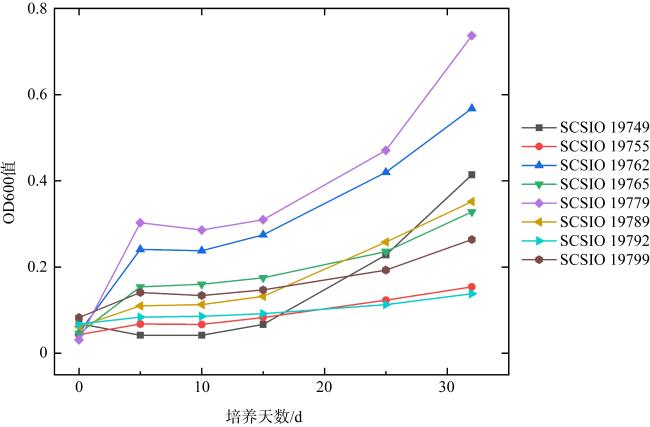
表4 潜在EHMC降解菌株信息Tab. 4 The information of potential EHMC-degrading strains |
| 菌株来源 | 菌株编号 | 种名 | 相似度/% |
|---|---|---|---|
| 杯形珊瑚 | SCSIO 19749 | Bacillus altitudinis 41KF2b | 100.00 |
| 角蜂巢珊瑚 | SCSIO 19792 | Brevundimonas diminuta ATCC 11568 | 99.77 |
| 角蜂巢珊瑚 | SCSIO 19789 | Paracoccus rhizosphaerae CC-CCM15-8 | 99.17 |
| 鹿角珊瑚 | SCSIO 19765 | Acuticoccus sediminis PTG4-2 | 100.00 |
| 鹿角珊瑚 | SCSIO 19799 | Phycicoccus jejuensis NRRL B-24460 | 99.13 |
| 鹿角珊瑚 | SCSIO 19755 | Rhodococcus hoagii DSM 20295 | 99.42 |
| 牡丹珊瑚 | SCSIO 19779 | Mycolicibacterium poriferae ATCC 35087 | 99.86 |
| 滨珊瑚 | SCSIO 19762 | Paracoccus aerius 011410 | 97.59 |
| [1] |
陈飚, 2021. 南海珊瑚微生物组的空间变化及其环境适应机制[D]. 南宁: 广西大学.
|
| [2] |
崔倩倩, 刘朝阳, 2020. 石油烃类污染物的微生物修复研究进展[J]. 江西科学, 38(3): 326-330, 384.
|
| [3] |
李明, 2022. 南海广布种澄黄滨珊瑚和低纬度优势种疣状杯形珊瑚的群体遗传学研究[D]. 南宁: 广西大学.
|
| [4] |
李汶璐, 王志超, 杨文焕, 等, 2022. 微塑料对沉积物细菌群落组成和多样性的影响[J]. 环境科学, 43(5): 2606-2613.
|
| [5] |
钱程, 2022. 新型二苯甲酮-3脱毒降解菌Pandoraea pnomenusa的筛选及降解特性研究[D]. 上海: 上海师范大学.
|
| [6] |
田蕴, 郑天凌, 2004. 海洋环境中降解多环芳烃的微生物[J]. 海洋科学, 28(9): 50-55.
|
| [7] |
王任, 吴鸳鸯, 程巧鸳, 2023. 2021年市售防晒类化妆品中防晒剂使用情况分析[J]. 香料香精化妆品, (1): 35-41.
|
| [8] |
吴钟解, 陈石泉, 陈敏, 等, 2013. 海南岛造礁石珊瑚资源初步调查与分析[J]. 海洋湖沼通报, (2): 44-50.
|
| [9] |
赵美霞, 余克服, 张乔民, 等, 2008. 三亚鹿回头石珊瑚物种多样性的空间分布[J]. 生态学报, 28(4): 1419-1428.
|
| [10] |
周楠, 姜成英, 刘双江, 2016. 从环境中分离培养微生物: 培养基营养水平至关重要[J]. 微生物学通报, 43(5): 1075-1081.
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}