
浙南海岸带人工秋茄(Kandelia obovata)红树林与互花米草(Spartina alterniflora)盐沼土壤碳汇对比研究
|
吴雪(1993—), 女, 研究生, 研究方向为海洋生态学。email: 13478659432@163.com |
Copy editor: 孙翠慈
收稿日期: 2024-03-07
修回日期: 2024-04-17
网络出版日期: 2024-05-07
基金资助
国家质量基础设施体系专项海洋生态监测 NQI 研究成果集成示范应用(2023YFF0611804)
浙江省自然资源厅科技计划项目(2022-42)
浙江省领雁研发攻关计划项目(2022C03G1253994)
Comparative study on soil carbon sinks of artificial Kandelia obovate mangrove and Spartina alterniflora salt marsh in the southern Zhejiang coastal zone
Copy editor: SUN Cuici
Received date: 2024-03-07
Revised date: 2024-04-17
Online published: 2024-05-07
Supported by
Integrated Demonstration Application of NQI Research Results in Marine Ecological Monitoring of Research Program of National Quality Infrastructure System(2023YFF0611804)
Science and Technology Planning Project of Zhejiang Provincial Department of Natural Resources(2022-42)
"Leading Goose" R&D Program of Zhejiang(2022C03G1253994)
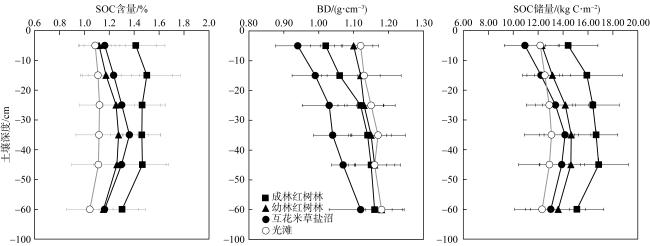
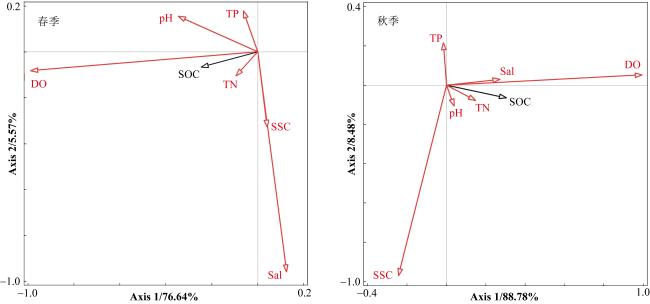
滨海湿地修复是提升海岸带蓝碳生态系统碳汇功能的重要举措。浙江省持续开展了大规模的红树林引种造林工程, 然而, 人工种植红树林后湿地固碳能力及其影响因素尚不清晰。本研究以乐清湾人工种植红树林(秋茄Kandelia obovata)为对象, 研究了不同林龄土壤有机碳含量、单位面积碳储量及其垂直分布特征, 并与互花米草(Spartina alterniflora)盐沼和淤泥质光滩进行比较。结果表明,成林红树林土壤有机碳含量和储量最高, 且红树林和盐沼生境土壤有机碳含量和储量均显著高于光滩。互花米草盐沼土壤有机碳含量高于幼林红树林, 但由于互花米草盐沼土壤容重显著低于幼林红树林, 因此, 幼林红树林土壤有机碳储量略高于互花米草盐沼。不同生境土壤有机碳含量和储量都呈现随土壤深度先升高后下降的分布特征, 一般在20~50cm深度最高。采样区域和生境类型差异对土壤有机碳含量和储量影响显著, 并有显著的交互作用。孔隙水溶解氧、土壤总氮、土壤pH和孔隙水盐度与土壤有机碳含量呈显著正相关关系。本研究结果可为评估浙江省红树林北移后的固碳潜力以及湿地功能提升提供科学依据与数据支撑。
吴雪 , 赵鑫 , 辜伟芳 , 朱科桦 , 葛振鸣 . 浙南海岸带人工秋茄(Kandelia obovata)红树林与互花米草(Spartina alterniflora)盐沼土壤碳汇对比研究[J]. 热带海洋学报, 2025 , 44(1) : 172 -181 . DOI: 10.11978/2024052
Coastal wetland restoration is an important measure to improve the blue carbon function in the coastal zone. Zhejiang Province has continuously carried out a series of large-scale mangrove introduction and afforestation projects. Yet, the carbon sink capacity and influence of factors for the high-latitude artificial mangroves still remain unclear. In this study, the soil organic carbon (SOC) content, organic carbon stock per unit area and their vertical distribution characteristics of the Kandelia obovata forests with different ages were investigated. The SOC content and stock in the mangroves were also compared with the Spartina alterniflora salt marsh and bare mudflat. The results showed that the SOC content and stock were the highest in the adult mangroves. The SOC content and stock in both mangroves and salt marshes were significantly higher than those in the mudflat. The SOC content in salt marshes was higher than that in the young mangroves, but due to the significant lower bulk density of Spartina alterniflora salt marsh soils compared to young mangrove forests, the SOC stock in the young mangroves was slightly higher than that of salt marshes. The SOC content and stock in the different habitats showed the distribution characteristics of first increasing and then decreasing with the soil depth, reaching the highest values at the soil depth of 20~50 cm. Site location and habitat type had a significant relationship with SOC content and stock, with significant interaction between site location and habitat type. Dissolved oxygen in pore water, soil total nitrogen, soil pH and pore water salinity were significantly associated with the SOC content. The results of this study can provide scientific basis and data support for assessing the carbon sequestration potential of artificial mangrove forests after northward migration, as well as wetland protection and restoration strategy.
Key words: coastal wetland; mangrove; exotic plants; soil organic carbon; carbon stock
图1 浙江省乐清湾滨海湿地红树林、互花米草盐沼和光滩土壤碳汇采样区域(包括茅坦岛、江岩岛、大青岛、茅埏岛和太平塘)左图基于国家测绘地理信息局标准地图服务网站下载的审图号为GS(2019)3333号的标准地图制作 Fig. 1 Sampling sites for soil carbon sink in the adult mangrove forests, junior mangrove forests, Spartina salt marsh and mudflats in the Yueqing Bay of the Zhejiang Province (including Maotan Island, Jiangyan Island, Daqing Island, Maoyan Island and Taipingtang) |
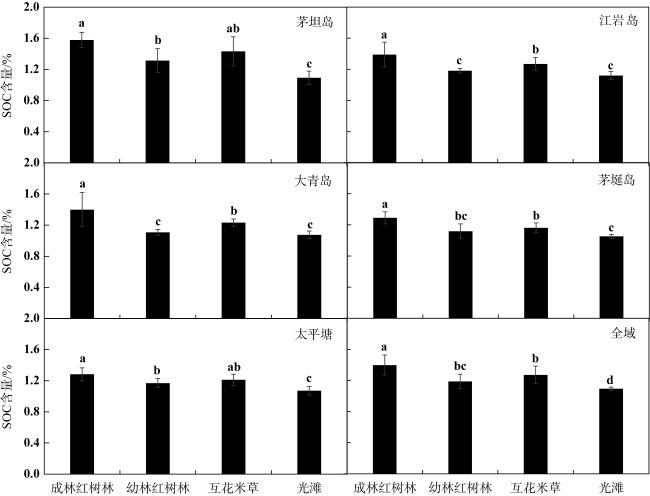
图2 茅坦岛、江岩岛、大青岛、茅埏岛和太平塘不同生境表层SOC含量(平均值±标准差)数据柱上的字母相同表示差异不显著, 字母不同则表示差异达到显著性水平(P<0.05, 方差分析) Fig. 2 The SOC content in the Maotan island, Jiangyan island, Daqing island, Maoyan island and Taipingtang (mean±SD). Different letters on the quantitative bars indicate significant differences (P<0.05, ANOVA) |
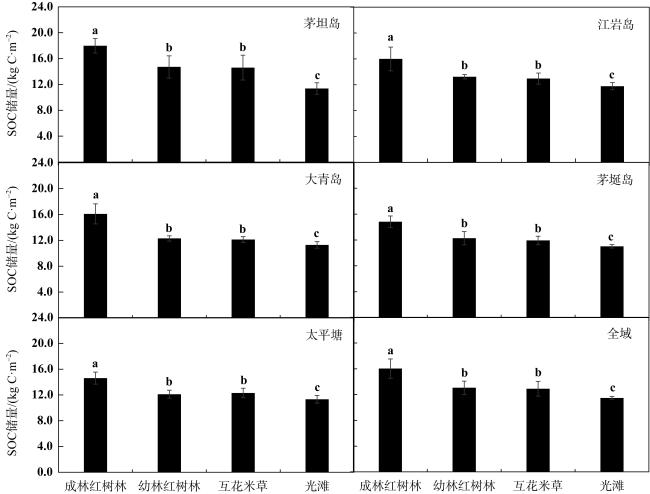
图3 茅坦岛、江岩岛、大青岛、茅埏岛和太平塘不同生境SOC储量(平均值±标准差)数据柱上的字母相同表示差异不显著, 字母不同则表示差异达到显著性水平(P<0.05, 方差分析) Fig. 3 The SOC stock in the Maotan island, Jiangyan island, Daqing island, Maoyan island and Taipingtang (mean±SD). Different letters on the quantitative bars indicate significant differences (P<0.05, ANOVA) |
表1. 季节、区域和生境类型对土壤表层SOC含量与储量的主效应与交互作用分析(F值与P值)Tab.1 Main and interactive effects (F and P values) of season, site and habitat type on the SOC content and stock |
| 主效应/交互作用组别 | df | F(SOC含量) | F(SOC储量) |
|---|---|---|---|
| 季节 | 1 | 0.02ns | 0.324ns |
| 区域 | 4 | 7.024** | 27.448** |
| 类型 | 3 | 41.480** | 35.406** |
| 季节×区域 | 4 | 1.205ns | 1.340ns |
| 类型×季节 | 3 | 2.882* | 2.763* |
| 区域×类型 | 12 | 2.699* | 3.109* |
| 季节×区域×类型 | 12 | 1.346ns | 1.253ns |
注: *表示 P<0.05, ** 表示P<0.01, ns表示 P>0.05 |
表2. 不同生境的土壤和孔隙水环境因子(平均值±标准差)Tab 2. Soil and pore water environmental variables in the different habitats (mean±SD) |
| 季节 | 生境类型 | pH (无量纲) | DO/(mg·L-1) | Sal/‰ | SSC/(g·kg-1) | TN/(g·kg-1) | TP/(mg·kg-1) |
|---|---|---|---|---|---|---|---|
| 春季 | 成林红树林 | 7.66±0.15a | 3.31±1.50b | 21.82±2.99a | 29.63±6.79a | 1.07±0.08ab | 688.82±25.46a |
| 幼林红树林 | 7.65±0.22a | 4.72±1.53a | 21.69±4.49a | 26.33±2.89a | 1.10±0.13ab | 699.64±38.15a | |
| 互花米草盐沼 | 7.64±0.32a | 3.51±1.88b | 22.79±4.22a | 27.15±4.68a | 1.17±0.09a | 700.55±36.49a | |
| 光滩 | 7.59±0.24a | 3.36±2.11b | 23.54±2.47a | 26.86±4.50a | 1.03±0.09b | 701.26±27.07a | |
| 秋季 | 成林红树林 | 7.33±0.18a | 4.28±2.54b | 29.42±1.87a | 37.67±13.57a | 1.18±0.15a | 638.70±12.64a |
| 幼林红树林 | 7.55±0.24a | 5.25±2.53a | 28.76±2.39a | 34.73±10.50a | 1.05±0.07ab | 632.82±11.98a | |
| 互花米草盐沼 | 7.29±0.18a | 2.63±2.50c | 29.72±3.97a | 33.34±16.63a | 1.08±0.09ab | 650.79±43.09a | |
| 光滩 | 7.49±0.24a | 4.50±2.84b | 29.66±1.26a | 33.56±11.93a | 0.99±0.06b | 629.80±14.05a |
注: 不同字母表示变量差异性在生境类型间达到显著性水平(P<0.05) |
| [1] |
陈桂香, 高灯州, 陈刚, 等, 2017. 互花米草入侵对我国红树林湿地土壤碳组分的影响[J]. 水土保持学报, 31(6): 249-256.
|
| [2] |
陈卉, 2013. 中国两种亚热带红树林生态系统的碳固定、掉落物分解及其同化过程[D]. 厦门: 厦门大学.
|
| [3] |
陈秋夏, 杨升, 王金旺, 等, 2019. 浙江红树林发展历程及探讨[J]. 浙江农业科学, 60(7): 1177-1181 (in Chinese).
|
| [4] |
胡雪红, 张立, 周炎武, 等, 2020. 我国滨海湿地生态修复领域规范的现状与分析[J]. 热带海洋学报, 39(6): 131-139.
|
| [5] |
刘景双, 杨继松, 于君宝, 等, 2003. 三江平原沼泽湿地土壤有机碳的垂直分布特征研究[J]. 水土保持学报, 17(3): 5-8.
|
| [6] |
逄柏鹏, 2012. 九龙江口不同恢复阶段秋茄红树林底质有机碳储量及其来源的研究[D]. 厦门: 厦门大学.
|
| [7] |
唐剑武, 叶属峰, 陈雪初, 等, 2018. 海岸带蓝碳的科学概念、研究方法以及在生态恢复中的应用[J]. 中国科学: 地球科学, 48(6): 661-670.
|
| [8] |
张天雨, 葛振鸣, 张利权, 等, 2015. 崇明东滩湿地植被类型和沉积特征对土壤碳、氮分布的影响[J]. 环境科学学报, 35(3): 836-843.
|
| [9] |
赵彩云, 李俊生, 赵相健, 等, 2015. 中国沿海互花米草入侵与防控管理[M]. 北京: 科学出版社.
|
| [10] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员, 2007. 海洋监测规范第4部分:海水分析GB17378. 4-2007 [S]. 北京: 中国标准出版社: 92-95.
General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, Standardization Administration of the People’s Republic of China, 2007. The specification for marine monitoring—Part 4: Seawater analysis GB17378. 4-2007 [S]. Beijing: Standards Press of China: 92-95 (in Chinese).
|
| [11] |
中华人民共和国农业部, 2006. 土壤检测第16部分:土壤水溶性盐总量的测定 NT/Y1121. 16-2006 [S]. 北京: 中国农业出版社: 1-2.
Ministry of Agriculture of the People’s Republic of China, 2006. Soil Testing—Part 16: Method for determination of total water-soluble salt NT/Y1121. 16-2006 [S]. Beijing: China Agriculture Press: 1-2 (in Chinese).
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
World Meteorological Organization (WMO), (2022-10-26)[2024-03-07]. The state of greenhouse gases in the atmosphere based on global observations through 2021, World Meteorological Organization[R/OL]. WMO Greenhouse Gas Bulletin, 18. https://library.wmo.int/idurl/4/58743.
|
| [40] |
|
| [41] |
|
| [42] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}