
别样玫瑰变色杆菌(Aliiroseovarius sp.)Z3基因组测序及比较基因组分析
|
刘巍(1996—), 女, 甘肃省白银市人, 硕士研究生, 从事微生物生态学研究。email: |
Copy editor: 姚衍桃
收稿日期: 2021-02-13
修回日期: 2021-04-08
网络出版日期: 2021-04-12
基金资助
国家自然科学基金(31672658)
浙江省教育厅一般项目(Y201839299)
宁波市农业重大专项(2017C110001)
Draft genome sequence and comparative genome analysis of Alliroseovarius sp. Z3
Copy editor: YAO Yantao
Received date: 2021-02-13
Revised date: 2021-04-08
Online published: 2021-04-12
Supported by
National Natural Science Foundation of China(31672658)
General Research Project of Zhejiang Education Department, China(Y201839299)
Agricultural Major Project of Ningbo, China(2017C110001)
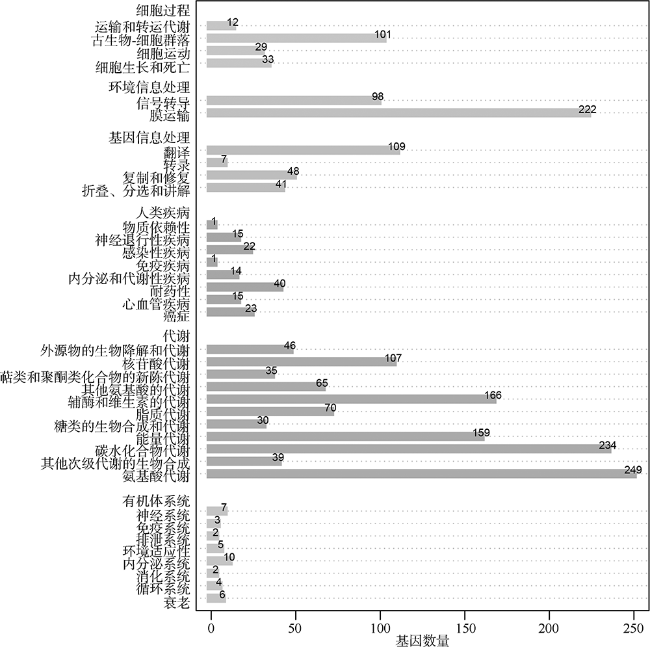
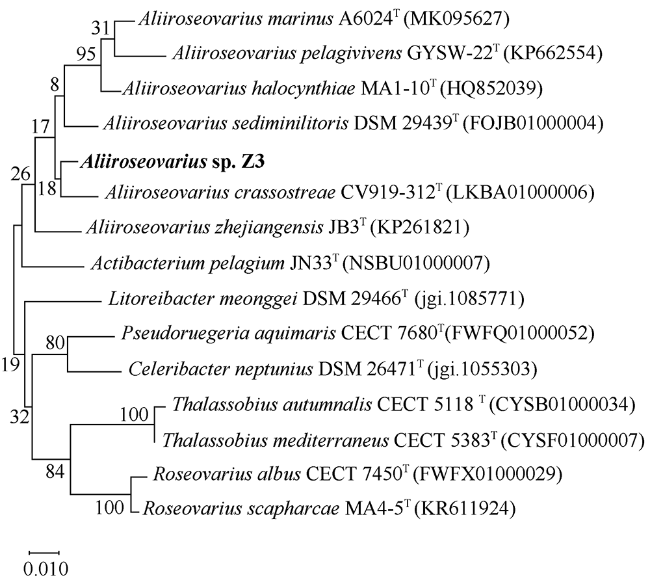
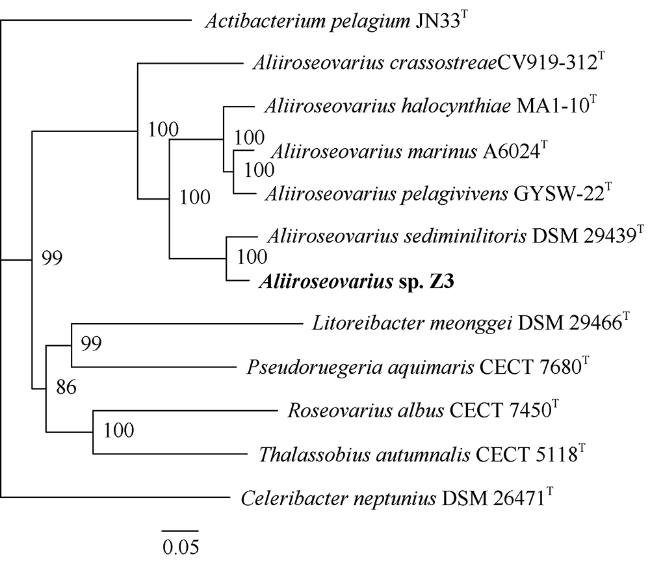
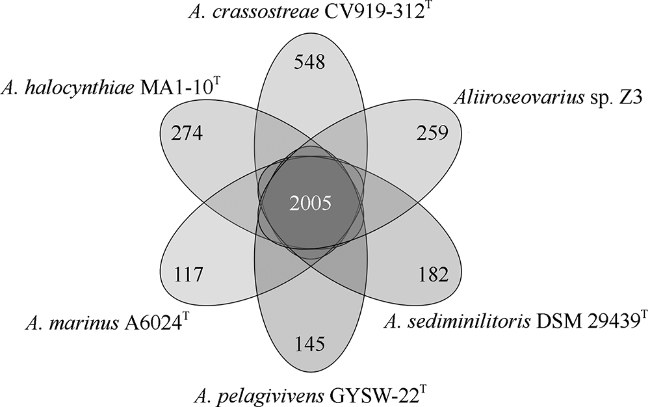
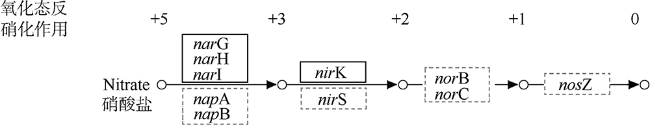
菌株Z3是一株从凡纳滨对虾(Litopenaeus vannamei)肠道样品中分离出的细菌。本研究利用Illumina HiSeq测序平台对其进行了测序, 用SOAPdenovo等软件进行基因组组装、系统发育分析、基因预测和功能注释, 并与别样玫瑰变色杆菌属(Aliiroseovarius)已知的5个种的模式株进行了比较基因组分析。基因组注释结果显示, Z3基因组大小为3525503bp, GC(guanylate and cytidylic acid)含量为59.4%, 预测包含3509个编码蛋白基因。通过16S rRNA基因全长序列比对、平均核苷酸一致性(ANI)、DNA-DNA杂交(DDH)和共线性分析发现, 菌株Z3与Aliiroseovarius crassostreae CV919-312T的16S rRNA基因全长序列相似度最高, 为98.20%, 与Aliiroseovarius sediminilitoris DSM 29439T的DDH和ANI值最高, 分别为26.80%和84.95%, 属于Alliroseovarius属的新种, 将其命名为Aliiroseovarius sp. Z3。经比较基因组分析发现, Aliiroseovarius sp. Z3与5个近缘的模式株细菌共享2005个直系同源核心基因簇; Z3拥有的413个独立基因经注释, 主要与碳水化合物转运与代谢、氨基酸转运与代谢、复制、重组及修复等功能相关。功能注释发现Z3具有进行完整的反硝化途径的相关基因, 其利用的可能是环境中的硝酸盐、亚硝酸盐等。本研究对Z3全基因序列的注释、功能和比较基因组的分析, 不仅丰富了Aliiroseovarius属细菌基因组资源, 还为深入研究其反硝化特性等提供了基础。
关键词: 对虾肠道; Aliiroseovarius sp. Z3; 系统发育分析; 反硝化; 比较基因组
刘巍 , 郭海朋 , 董鹏生 , 燕孟琛 , 张德民 . 别样玫瑰变色杆菌(Aliiroseovarius sp.)Z3基因组测序及比较基因组分析[J]. 热带海洋学报, 2022 , 41(1) : 52 -61 . DOI: 10.11978/2021018
Alliroseovarius sp. Z3 was isolated from guts of Litopenaeus vannamei. In the study, Illumina Hiseq sequencing platform was used for genome sequencing; then, the genome was assembled, annotated by software SOAPdenovo etc., and compared with other five similar type strains. Genome annotation results showed that Z3 genome consists of 3525503 bp with a guanine-cytosine (GC) content of 59.4%, and 3509 protein-coding genes. The results of 16S rRNA sequence alignment, average nucleotide identity (ANI), DNA-DNA hybridization (DDH) and collinearity analysis showed that strain Z3 had the highest 16S rRNA similarity with Aliiroseovarius crassostreae CV919-312T, at 98.20%, and had the highest DDH and ANI values with Aliiroseovarius sediminilitoris DSM 29439T, at 26.80% and 84.95%, respectively. It was considered to be a new species of Alliroseovarius, and was named Aliiroseovarius sp. Z3. Comparative genomic analysis displayed that Aliiroseovarius sp. Z3 shared 2005 core orthologous gene clusters with other five similar type strains, and Z3 had 413 specific genes that are related to carbohydrate transport and metabolism, amino acid transport and metabolism, replication, recombination and repair, etc. After functional annotations, we found that Z3 possesses a complete denitrification pathway, indicating that Z3 can utilize nitrate and nitrite in the environment. These results related to the genome features and comparative genome of strain Z3 not only enrich the gene source of Aliiroseovarius genus, but also provide a molecular basis for studying its characteristics of denitrification.
图3 基于最大似然法构建的别样玫瑰变色杆菌Z3 16S rRNA基因的系统发育树Fig. 3 The phylogenetic tree of Aliiroseovarius sp. Z3 16S rRNA gene based on the maximum likelihood method |
图4 基于单拷贝直系同源基因构建的系统发育树Fig. 4 The phylogenetic tree based on the single copy orthologous gene families |
表1 6株别样玫瑰变色杆菌属细菌平均核苷酸同一性的值Tab. 1 Average nucleotide identity (ANI) values of six Aliiroseovarius spp. |
| Aliiroseovarius sp. Z3 | A. crassostreae CV919-312T | A. halocynthiae MA1-10T | A. marinus A6024T | A. pelagivivens GYSW-22T | A. sediminilitoris DSM 29439T | |
|---|---|---|---|---|---|---|
| Aliiroseovarius sp. Z3 | * | 84.51% | 84.33% | 84.87% | 84.49% | 84.95% |
| A. crassostreae CV919-312T | 84.51% | * | 84.82% | 84.95% | 85.04% | 83.92% |
| A. halocynthiae MA1-10T | 84.33% | 84.81% | * | 85.82% | 85.52% | 83.69% |
| A. marinus A6024T | 84.87% | 84.94% | 85.83% | * | 86.50% | 83.62% |
| A. pelagivivens GYSW-22T | 84.51% | 85.02% | 85.52% | 86.50% | * | 83.63% |
| A. sediminilitoris DSM 29439T | 84.95% | 83.92% | 83.69% | 83.61% | 83.65% | * |
表2 别样玫瑰变色杆菌Z3与其他5株模式株DNA- DNA杂交的值Tab. 2 DNA-DNA hybridization (DDH) values between Aliiroseovarius sp. Z3 and other five Aliiroseovarius spp. |
| Aliiroseovarius sp. Z3 | |
|---|---|
| A. crassostreae CV919-312T | 20.90% |
| A. sediminilitoris DSM 29439T | 26.80% |
| A. marinus A6024T | 20.60% |
| A. halocynthiae MA1-10T | 20.30% |
| A. pelagivivens GYSW-22T | 20.30% |
表3 Z3和与其近缘的5株别样玫瑰变色杆菌属模式株的基因组基本特征Tab. 3 Genome features summary of Z3 and other five similar type strains from Aliiroseovarius genus |
| 名称 | 大小/Mb | GC含量/% | 编码蛋白 | 分离来源 |
|---|---|---|---|---|
| Aliiroseovarius sp. Z3 | 3.53 | 59.4 | 3509 | 中国浙江宁波: 凡纳滨对虾肠道 |
| A. crassostreae CV919-312T | 3.73 | 58.4 | 3693 | 美国缅因州达马里斯科塔河: 患病牡蛎 |
| A. sediminilitoris DSM 29439T | 3.41 | 58.7 | 3323 | 中国南部吉野岛: 近海沉积物 |
| A. marinus A6024T | 3.13 | 59.9 | 3027 | 中国山东日照: 海水 |
| A. halocynthiae MA1-10T | 3.39 | 57.1 | 3349 | 南海: 海鞘 |
| A. pelagivivens GYSW-22T | 3.33 | 58.1 | 3218 | 韩国巨济岛: 海水 |
图5 菌株Z3与其他5个别样玫瑰变色杆菌Aliiroseovarius crassostreae CV919-312T(a)、Aliiroseovarius sediminilitoris DSM 29439T(b)、Aliiroseovarius halocynthiae MA1-10T(c)、Aliiroseovarius marinus A6024T(d)和Aliiroseovarius pelagivivens GYSW-22T(e)的共线性分析图中数值表示基因片段大小 Fig. 5 Synteny blocks between Aliiroseovarius sp. Z3 and other five Aliiroseovarius spp., Aliiroseovarius. crassostreae CV919-312T (a), Aliiroseovarius sediminilitoris DSM 29439T (b), Aliiroseovarius. halocynthiae MA1-10T (c), Aliiroseovarius marinus A6024T (d), and Aliiroseovarius pelagivivens GYSW-22T (e). The value in the figure indicates the size of the gene fragment |
图6 6株别样玫瑰变色杆菌的直系同源基因和特异性基因簇的数量Fig. 6 The numbers of orthologous and unique gene clusters among six Aliiroseovarius spp. |
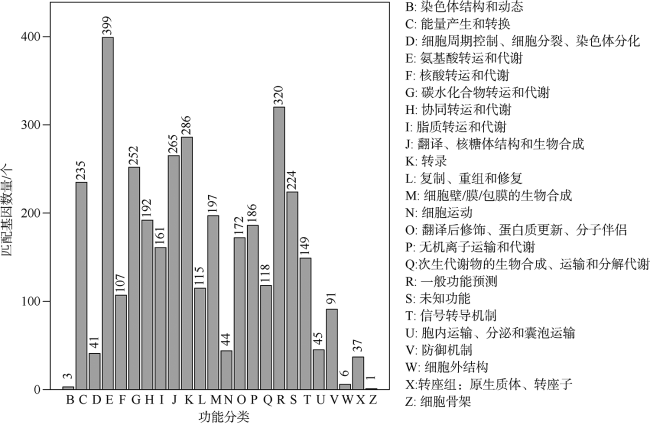
表4 基于COG数据库比对的6株别样玫瑰变色杆菌属细菌的直系同源核心基因和Z3的特异性基因Tab. 4 The number of core orthologous gene clusters of six Aliiroseovarius spp. and specific 418 genes in the genome of Aliiroseovarius sp. Z3 based on COG categories analysis |
| COG分类 | 核心基因/个 | Z3的特异性基因/个 |
|---|---|---|
| 未分类(Unassigned) | 992 | 220 |
| 能量产生与转换(C) | 149 | 13 |
| 细胞周期调控、细胞分裂、染色体分区(D) | 9 | 2 |
| 氨基酸转运与代谢(E) | 305 | 14 |
| 核苷酸转运与代谢(F) | 22 | 2 |
| 碳水化合物的转运与代谢(G) | 139 | 15 |
| 辅酶转运与代谢(H) | 97 | 6 |
| 脂类转运与代谢(I) | 78 | 6 |
| 翻译、核糖体结构和生物合成(J) | 30 | 3 |
| 转录(K) | 217 | 12 |
| 复制、重组和修复(L) | 31 | 12 |
| 细胞壁/膜/包体生物合成(M) | 76 | 10 |
| 细胞运动(N) | 2 | 0 |
| 翻译后修饰、蛋白质周转和分子伴侣(O) | 46 | 8 |
| 无机离子转运与代谢(P) | 235 | 12 |
| 次生代谢物的生物合成、转运和分解代谢(Q) | 48 | 2 |
| 功能未知(S) | 184 | 68 |
| 信号转导机制(T) | 110 | 4 |
| 细胞内运输、分泌和囊泡运输(U) | 17 | 3 |
| 防御机制(V) | 44 | 6 |
| [1] |
刘志迎, 许海, 詹旭, 等, 2019. 蓝藻水华对太湖水柱反硝化作用的影响[J]. 环境科学, 40(3): 1261-1269.
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}