
黄鹂无齿鲹的驯养及人工繁殖技术研究*
*感谢莫洋、谢又兴、丁运源、关善尹等为试验提供的帮助。
|
施钢(1978—), 男, 江苏省海门区人, 高级工程师, 从事鱼类繁殖生物学研究。email: shign@126.com |
Copy editor: 殷波 , YIN Bo
收稿日期: 2024-04-26
修回日期: 2024-07-01
网络出版日期: 2024-07-19
基金资助
广东海洋大学创新强校工程项目(2019KTSCS056)
Study on the domestication and artificial propagation technology of golden trevally, Gnathanodon speciosus*
Copy editor: YIN Bo
Received date: 2024-04-26
Revised date: 2024-07-01
Online published: 2024-07-19
Supported by
Guangdong Ocean University “Innovate and strengthen the school project”(2019KTSCS056)
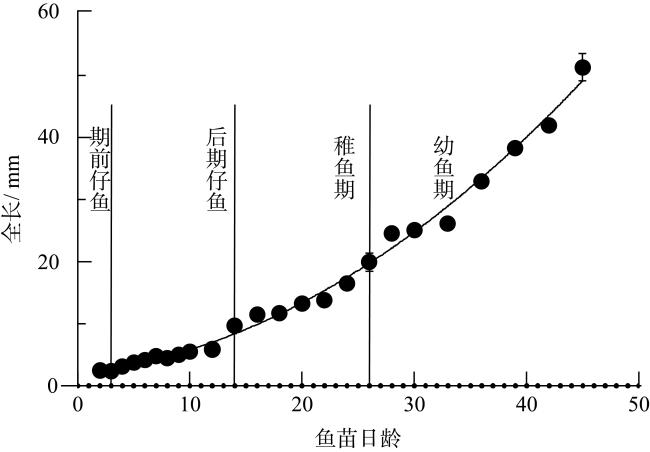
2017年至2023年对黄鹂无齿鲹野生群体进行驯养、催熟, 并对其人工繁殖技术进行了初步研究。自然成熟产卵的受精卵为透明、球形浮性卵, 卵黄囊有油球1个, 卵径(0.85 ± 0.04)mm, 油球径(0.23 ± 0.02)mm, 每克卵量(3197 ± 210)粒, 受精卵在水温(28 ± 0.5)℃、盐度30‰、pH 8.0~8.2、溶解氧6.05~7.13mg·L-1的海水中, 胚胎历时17h17min完成胚胎发育, 其胚胎发育分为卵裂期、囊胚期、原肠胚期、神经胚期、器官形成期和孵化期6个阶段。孵化的生态学总积温为483.84℃·h, 初孵仔鱼全长(2.01 ± 0.14)mm, 受精卵孵化率为94.95%; 在水温(29.5 ± 2.0)℃、盐度26‰~32‰、pH 7.8~9.0、溶解氧4.90~ 7.50mg∙L-1的海水中, 完成了前期仔鱼、后期仔鱼、稚鱼期、幼鱼期共4个阶段培育, 从初孵仔鱼期至前期仔鱼期成活率为91.49%, 出膜后14日龄进入稚鱼期, 出膜后26日龄完成变态成为幼鱼, 出膜后45日龄幼鱼体色和形态已与成鱼基本一致, 全长(51.05 ± 4.81)mm。本研究完成了黄鹂无齿鲹的苗种培育, 总育苗成活率32.62%, 并可进行全人工繁殖。
施钢 , 黄晓晴 , 曾珍 , 孙琛琛 , 潘传豪 . 黄鹂无齿鲹的驯养及人工繁殖技术研究*[J]. 热带海洋学报, 2025 , 44(2) : 115 -123 . DOI: 10.11978/2024093
An artificially bred protocol, developed for wild Gnathanodon speciosus, also named as golden trevally, was presented in this study. The embryonic development and growth process of the fish was also introduced. The wild fish were domesticated from 2017 to 2023. The fertilized eggs laid by naturally matured fish were transparent and buoyant. Single oil globule ((0.23 ± 0.02) mm in diameter) was observed in the yolk sac of eggs ((0.85 ± 0.04) mm in diameter). The number of eggs per gram was (3197 ± 210). The embryonic development went through six stages: cleavage, blastocyst, gastrulation, neuroblast, organogenesis and hatching. The hatching of fertilized eggs took 17 h 17 min and the hatching rate was 94.95% in sea water (salinity 30‰, pH 8.0 ~ 8.2, DO 6.05 ~ 7.13 mg∙L-1) at (18 ± 0.5) ℃. The ecological accumulated temperature was 483.84 ℃·h. The body length of newly hatched larvae were (2.01 ± 0.14) mm. The nurture of fish fingerlings went through four stages: early larval, late larval, juvenile and fingerling, and took 45 dph with a 32.62% survival rate in sea water (salinity 26‰~32‰, pH 7.8~9.0, DO 4.90~7.50 mg∙L-1) at (29.5 ± 2.0)℃. The transformation from juvenile to fingerling fish was completed at 26 dph. At 45 dph of growth, the fingerling fish ((51.05 ± 4.81) mm in body length) were similar to adult fish in body shape and color. This study indicated that the golden trevally grew and spawned naturally in artificial bred condition and could be artificially reproduced.
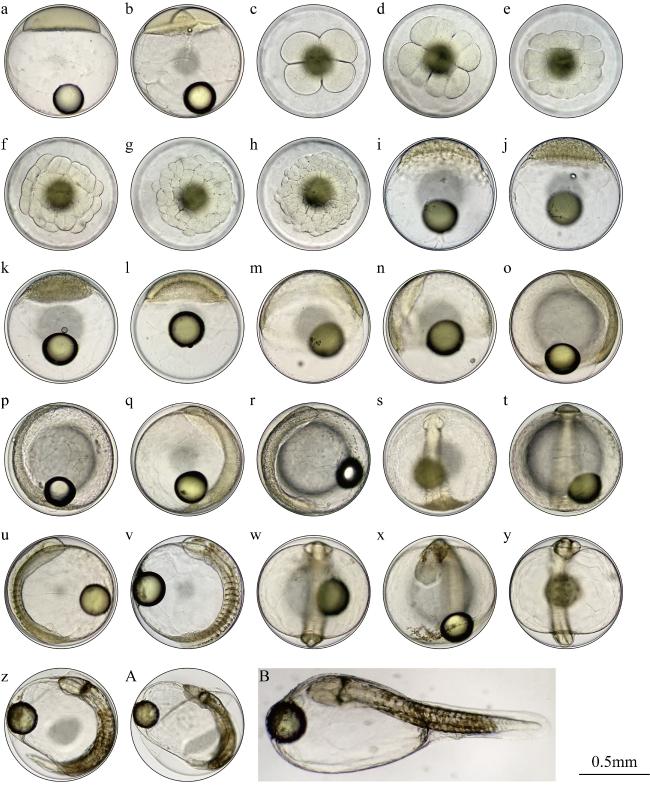
图1 黄鹂无齿鲹胚胎发育图a. 胚盘隆起期; b. 2细胞期; c. 4细胞期; d. 8细胞期; e. 16细胞期; f. 32细胞期; g. 64细胞期; h. 多细胞期; i. 桑葚期; j. 高囊胚期; k. 低囊胚期; l. 原肠早期; m. 原肠中期; n. 原肠晚期; o. 胚体形成期; p. 胚孔封闭期; q. 视囊形成期; r. 肌节出现期; s. 听囊形成期; t. 脑泡形成期; u. 心脏形成期(Ⅰ); v. 心脏形成期(Ⅱ); w. 尾芽期; x. 晶体形成期; y. 心脏跳动期; z. 将孵期; A. 孵化期; B. 初孵仔鱼 Fig. 1 Diagram of embryonic development of G. speciosus. a. Blastodisc formation; b. 2-cell stage; c. 4-cell stage; d. 8-cell stage; e. 16-cell stage; f. 32-cell stage; g. 64-cell stage; h. multi-cell stage; i. blackberry period; j. high blastula stage; k. low blastula stage; l. early gastrula stage; m. middle gastrula stage; n. late gastrula stage; o. embryo body stage; p. closure of blastopore; q. optic vesicle stage; r. muscle burl stage; s. otocyst stage; t. brain vesicle stage; u. heart stage Ⅰ; v. heart stage Ⅱ; w. tail-bud stage; x. eye lens formation; y. heart beating stage; z. pre-hatching stage; A. hatching stage; B. newly-hatched larvae |
表1 黄鹂无齿鲹胚胎发育过程Tab. 1 Embryonic development of G. speciosus |
| 发育时相 | 发育日龄 | 主要特征 |
|---|---|---|
| 胚盘隆起期 | 0h 18min | 胚盘扩大隆起达到最大高度, 侧面观呈帽状(图1a) |
| 2细胞期 | 0h 30min | 第1次分裂, 胚盘形成2个对等细胞(图1b) |
| 4细胞期 | 0h 45min | 第2次分裂, 胚盘形成4个细胞(图1c) |
| 8细胞期 | 0h 49min | 第3次分裂, 形成8个形态大小略有差异的细胞(图1d) |
| 16细胞期 | 1h 04min | 第4次分裂, 形成16个细胞(图1e) |
| 32细胞期 | 1h 17min | 第5次分裂, 形成32个细胞(1f) |
| 64细胞期 | 1h 27min | 第6次分裂, 形成64个细胞, 细胞变小并开始重叠(图1g) |
| 多细胞期 | 1h 42min | 细胞持续分裂, 体积变小, 数量增多, 呈多细胞胚体(图1h) |
| 桑葚期 | 1h 54min | 持续分裂, 细胞变得更小, 细胞团与桑葚球相似(图1i) |
| 高囊胚期 | 2h 17min | 细胞集中且界限不明显, 胚盘隆起, 侧面观呈高帽状(图1j) |
| 低囊胚期 | 2h 48min | 胚盘逐渐降低, 胚盘边缘细胞向植物极下包(图1k) |
| 原肠早期 | 4h 34min | 胚盘边缘细胞增多并下包, 背面可见内卷形成的胚环(图1l) |
| 原肠中期 | 6h 01min | 胚层细胞下包达卵黄1/3以上, 胚环更明显, 形成胚盾雏形(图1m) |
| 原肠晚期 | 6h 42min | 胚层细胞下包达卵黄1/2~2/3, 胚盾明显延长(图1n) |
| 胚体形成期 | 7h 05min | 胚体背面增厚, 形成神经板, 胚体轮廓清晰(图1o) |
| 胚孔封闭期 | 7h 50min | 胚层下包完全, 胚孔封闭(图1p) |
| 视囊形成期 | 8h 34min | 胚体头部两侧出现突出的视囊, 中部有模糊的肌节2节(图1q) |
| 肌节出现期 | 8h 54min | 眼囊中央出现一凹陷, 视杯逐渐形成, 肌节3~4对(图1r) |
| 听囊形成期 | 9h 36min | 胚体在视囊后形成比视囊稍小的听囊1对, 肌节5~6对(图1s) |
| 脑泡形成期 | 10h 14min | 两视囊中间位置出现板状脑泡, 肌节7~8对(图1t) |
| 心脏形成期 | 11h 05min | 心脏、脊索轮廓清晰, 肌节10~13对(图1u、图1v) |
| 尾芽期 | 12h 42min | 胚体后端突出形成尾芽并与卵黄囊分离, 胚体鳍褶形成(图1w) |
| 晶体形成期 | 13h 23min | 视囊出现晶状体, 肌节18对以上(图1x) |
| 心脏跳动期 | 14h 10min | 心脏开始由慢至快自主搏动(图1y) |
| 将孵期 | 16h 11min | 胚体从偶尔到频繁抽动, 孵出前心跳频率每分钟70~80次(图1z) |
| 孵化期 | 17h 17min | 胚体破膜而出, 尾部不断摆动, 脱去卵膜(图1A) |
| 初孵仔鱼 | 17h 17min | 仔鱼(图1B) |
图2 黄鹂无齿鲹仔稚幼鱼发育图a. 出膜后1日龄; b. 出膜后2日龄; c. 出膜后3日龄; d. 出膜后4日龄; e. 出膜后5日龄; f. 出膜后6日龄; g. 出膜后7日龄; h. 出膜后8日龄; i. 出膜后10日龄; j. 出膜后12日龄; k. 出膜后14日龄; l. 出膜后16日龄; m. 出膜后18日龄; n. 出膜后22日龄; o. 出膜后24日龄; p. 出膜后26日龄; q. 出膜后28日龄; r. 出膜后30日龄; s. 出膜后36日龄; t. 出膜后42日龄; u. 出膜后45日龄 Fig. 2 larvae, juvenile stage and fingerling fish of G. speciosus. a. 1 dph; b. 2 dph; c. 3 dph; d. 4 dph; e. 5 dph; f. 6 dph; g. 7 dph; h. 8 dph; i. 10 dph; j. 12 dph; k. 14 dph juvenile stage; l. 16 dph juvenile stage; m. 18 dph juvenile stage; n. 22 dph juvenile stage; o. 24 dph juvenile stage; p. 26 dph fingerling fish; q. 28 dph fingerling fish; r. 30 dph fingerling fish; s. 36 dph fingerling fish; t. 42 dph fingerling fish; u. 45 dph fingerling fish |
| [1] |
陈伟洲, 许鼎盛, 王德强, 等, 2007. 卵形鲳鲹人工繁殖及育苗技术研究[J]. 台湾海峡, 26(3): 435-442.
|
| [2] |
冲山宗雄, 1988. 日本产稚鱼图鉴[M]. 东京: 东海大学出版会:ⅶ.
|
| [3] |
黄贤克, 单乐州, 闫茂仓, 等, 2017. 黄姑鱼胚胎发育及其与温度和盐度的关系[J]. 海洋科学, 41(7): 44-50.
|
| [4] |
黄炜坪, 甘松永, 王庆, 等, 2024. 黄鹂无齿鲹胚胎及胚后发育特征[J]. 水产学报, 48(4): 73-85.
|
| [5] |
邝杰华, 陈刚, 马骞, 等, 2021. 军曹鱼的胚胎发育及仔稚鱼形态观察[J]. 水产学报, 45(11): 1814-1824.
|
| [6] |
黎祖福, 陈省平, 庄余谋, 等, 2006. 鞍带石斑鱼人工繁殖与鱼苗培育技术研究[J]. 海洋水产研究, 27(3): 78-85.
|
| [7] |
区又君, 何永亮, 李加儿, 等, 2012. 卵形鲳鲹胚后发育阶段的体色变化和鳍的分化[J]. 热带海洋学报, 31(1): 62-66.
|
| [8] |
彭志东, 2007. 卵形鲳鲹的人工繁育技术[J]. 内陆水产, (9): 14-15 (in Chinese).
|
| [9] |
苏跃中, 郑智莺, 游岚, 等, 1997. 大黄鱼Pseudosciaena crocea (Richardson)人工繁殖及育苗技术的研究[J]. 现代渔业信息, 12(5): 21-27.
|
| [10] |
苏志星, 岳彦峰, 蒋科技, 等, 2021. 国内外海水观赏鱼产业与研究现状[J]. 南方水产科学, 17(1): 101-112.
|
| [11] |
孙典荣, 陈铮, 2013. 南海鱼类检索[M]. 北京: 海洋出版社.
|
| [12] |
万瑞景, 张仁斋, 2016. 中国近海及其邻近海域鱼卵与仔稚鱼[M]. 上海: 上海科学技术出版社: 205-209.
|
| [13] |
伍汉霖, 钟俊生, 2021. 中国海洋及河口鱼类系统检索[M]. 北京: 中国农业出版社.
|
| [14] |
杨龙, 李昂, 李步苏, 等, 2021. 鲹科鱼类DNA条形码分类系统[J]. 中国水产科学, 28(12): 1523-1535.
|
| [15] |
杨培民, 金广海, 刘义新, 等, 2022. 瓦氏雅罗鱼胚后发育与早期生长[J]. 水产科学, 41(3): 379-387.
|
| [16] |
詹炜, 楼宝, 陈睿毅, 等, 2016. 小黄鱼(Larimichthys polyactis)胚胎发育及仔、稚鱼形态特征观察[J]. 海洋与湖沼, 47(5): 1033-1039.
|
| [17] |
张海发, 刘晓春, 刘付永忠, 等, 2006. 斜带石斑鱼胚胎及仔稚幼鱼形态发育[J]. 中国水产科学, 13(5): 689-699.
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}