
基于微卫星标记9个缢蛏群体的遗传多样性分析
|
吴贵清(2000—), 男, 广西壮族自治区柳州市人, 硕士研究生, 从事贝类遗传与育种研究。email: 18077267931@163.com |
Copy editor: 殷波 , YIN Bo
收稿日期: 2024-06-01
修回日期: 2024-08-19
网络出版日期: 2024-08-30
基金资助
广西民族大学引进人才启动基金(2017KJQD007)
广西自然科学基金资助项目(2020GXNSFBA159010)
2022年广西壮族自治区大学生创新创业训练计划项目(S202210608162)
2022年广西壮族自治区大学生创新创业训练计划项目(S202210608163)
2023年广西壮族自治区大学生创新创业训练计划项目(S202310608040)
Analysis of genetic diversity in 9 populations of Sinonovacula constricta using microsatellite markers
Copy editor: YIN Bo
Received date: 2024-06-01
Revised date: 2024-08-19
Online published: 2024-08-30
Supported by
High Level Introduction of Talent Research Start-up Projects of Guangxi Minzu University(2017KJQD007)
Guangxi Natural Science Foundation Grant(2020GXNSFBA159010)
Innovation and Entrepreneurship Training Program for College Students of Guangxi in 2022(S202210608162)
Innovation and Entrepreneurship Training Program for College Students of Guangxi in 2022(S202210608163)
Innovation and Entrepreneurship Training Program for College Students of Guangxi in 2023(S202310608040)
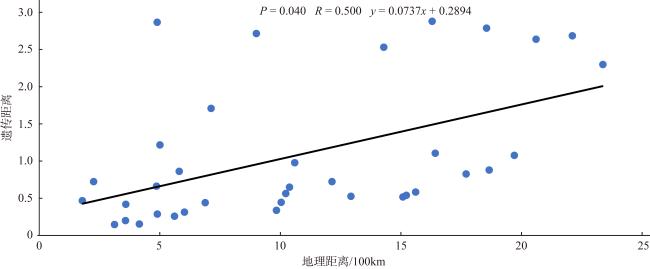
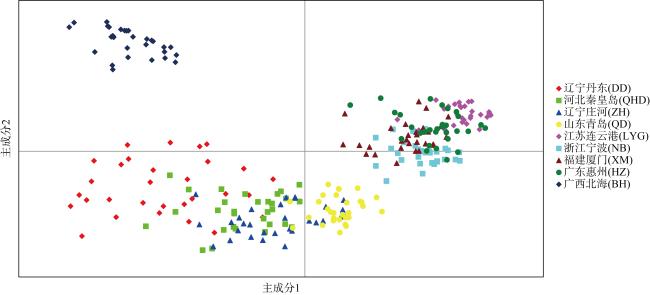
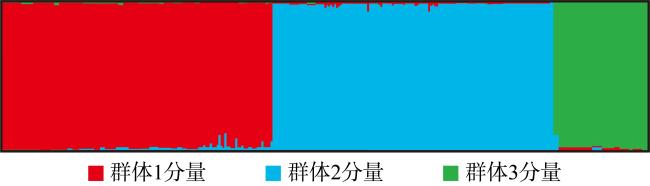
利用基于简化基因组测序(restriction site associated DNA sequencing, RAD-seq)技术开发了10对新的多态性微卫星引物, 对辽宁丹东(DD)、河北秦皇岛(QHD)、辽宁庄河(ZH)、山东青岛(QD)、江苏连云港(LYG)、浙江宁波(NB)、福建厦门(XM)、广东惠州(HZ)和广西北海(BH)共9个缢蛏群体进行了遗传多样性分析, 在270个缢蛏个体中共检测到了352个等位基因。平均等位基因数(Na, number of alleles)在3.2000~4.3000之间、平均有效等位基因数(Ne, effective number of alleles)在1.8789~2.5433之间; 观测杂合度(Ho, observed heterozygosity)范围为0.0000~0.9667、平均观测杂合度在0.3088~0.5533之间; 期望杂合度(He, expected heterozygosity)范围为0.0000~0.7945、平均期望杂合度在0.3456~0.5715之间; 平均多态信息含量(PIC, polymorphic information content)在0.3373~0.5989之间。遗传多样性评估显示, 9个缢蛏群体的遗传多样性水平属于中等。群体间的遗传分化系数(Fst, genetic differentiation coefficient)在0.0547~0.3511之间, 其中QHD和ZH群体之间的Fst值最低为0.0547, 而LYG和BH群体之间的Fst值最高为0.3511。基因流(Nm, gene flow)在0.4620至4.3204之间, 其中QHD和ZH群体之间的Nm值最高为4.3204, 而LYG和BH群体之间的Nm值最低为0.4620。分子方差分析(analysis of molecular variance, AMOVA)分析结果显示, 群体间的遗传变异占总变异的33.04% (p < 0.01), 而群体内的遗传变异占总变异的66.96% (p < 0.01), 表明遗传变异不仅存在于个体间, 也存在于群体间, 但个体间的遗传变异大于群体间的遗传变异。非加权组平均法(unweighted pair-group method with arithmetic means, UPGMA)、Structure软件的聚类结果以及主坐标分析(principal coordinate analysis, PCoA)结果一致。研究结果显示, 9个缢蛏群体可分为3大支: 第一支由QHD、ZH、QD、DD 4个群体组成; 第二支由LYG、NB、XM、HZ 4个群体组成; 第三支由BH群体独立成支。
吴贵清 , 李瑞华 , 肖意豪 , 陈彦林 , 罗璇 , 刘相全 , 朱佳杰 , 吴雪萍 . 基于微卫星标记9个缢蛏群体的遗传多样性分析[J]. 热带海洋学报, 2025 , 44(2) : 124 -136 . DOI: 10.11978/2024117
RAD-seq technology was utilized to develop 10 pairs of novel polymorphic microsatellite primers for assessing genetic 有时候diversity in nine populations of Sinonovacula constricta of Dandong in Liaoning (DD), Qinhuangdao in Hebei (QHD), Zhuanghe in Liaoning (ZH), Qindao in Shandong (QD), Lianyungang in Jiangsu (LYG), Ningbo in Zhejiang (NB), Xiamen in Fujian (XM), Huizhou in Guangdong (HZ), and Beihai in Guangxi (BH). In 270 individuals of S. constricta, a total of 352 alleles were detected. The average number of alleles (Na) ranged from 3.2000 to 4.3000, with the average effective number of alleles (Ne) falling between 1.8789 and 2.5433. Observed heterozygosity (Ho) varied from 0.0000 to 0.9667, with an average ranging from 0.3088 to 0.5533. Expected heterozygosity (He) spanned from 0.0000 to 0.7945, with an average of 0.3456 to 0.5715. The average polymorphic information content (PIC) ranged from 0.3373 to 0.5989. Genetic diversity analysis indicated a moderate level of genetic diversity across the nine populations of S. constricta. The genetic differentiation coefficient (Fst) among populations ranged from 0.0547 to 0.3511, with the lowest Fst value observed between QHD and ZH (0.0547) and the highest between LYG and BH (0.3511). Gene flow (Nm) values ranged from 0.4620 to 4.3204, with the highest Nm value recorded between QHD and QD (4.3204) and the lowest between LYG and BH (0.4620). AMOVA analysis revealed that 33.04% of the total genetic variation existed among populations (p < 0.01), while 66.96% was within populations (p < 0.01), indicating that the genetic variation was observed not only among individuals but also among populations. However, the variation among individuals was greater than that among populations. Consistent results were obtained from UPGMA clustering, Structure clustering, and principal coordinate analysis (PCoA). The study delineated three main groups among the nine populations: the first group comprised QHD, ZH, QD and DD populations; the second group included LYG, NB, XM, and HZ populations; and the third group was independently represented by BH population.
表1 9个缢蛏群体采样信息Tab. 1 Sampling information of 9 populations of S. constricta |
| 采样地点 | 经度 | 纬度 | 样本量/个 | 采样时间 |
|---|---|---|---|---|
| DD | 124°16′54′′E | 39°56′15′′N | 30 | 2022年11月22日 |
| QHD | 119°26′11′′E | 39°47′47′′N | 30 | 2022年11月05日 |
| ZH | 122°38′59′′E | 39°31′36′′N | 30 | 2022年10月23日 |
| QD | 120°67′16′′E | 36°16′06′′N | 30 | 2022年10月15日 |
| LYG | 119°17′08′′E | 34°46′50′′N | 30 | 2022年9月27日 |
| NB | 121°43′55′′E | 29°32′01′′N | 30 | 2022年9月18日 |
| XM | 118°11′50′′E | 24°29′18′′N | 30 | 2022年8月25日 |
| HZ | 114°38′26′′E | 22°45′40′′N | 30 | 2022年8月12日 |
| BH | 109°06′33′′E | 21°25′25′′N | 30 | 2022年7月10日 |
表2 10对缢蛏微卫星引物的信息Tab. 2 Information of 10 pairs of microsatellite locus of S. constricta |
| 位点 | 序列号 | 引物序列(5′-3′) | 退火温度/℃ | 片段大小/bp |
|---|---|---|---|---|
| SC03 | PP409554 | F: CTTATCTTTATCCTCCTCGCCAT R: TGATGATAATGTTGAGGCTGTTG | 54 | 120~160 |
| SC04 | PP409555 | F: AGATAAAGCCAGGAAAACACAGA R: GTTTACAATGAAAAGACAGATGGC | 52 | 110~150 |
| SC05 | PP409556 | F: TCTTGGTTAGAGTGGGTTGTTGT R: TGCTTTCTAATGTGCCTTTCATT | 52 | 130~170 |
| SC06 | PP409557 | F: TTTCATGTTGTTGGTTGTTTGTT R: AATGTTGCCAAATCAAAACATTC | 54 | 110~150 |
| SC07 | PP409558 | F: CATCATCATTAACCGTCCTCCTA R:AGAAATTGGCATTGATAGCAAAA | 52 | 120~160 |
| SC08 | PP409559 | F: CCATGCATGAGAGTTAAGTAACAAA R: CCAAGAATGCACAATAAAATTGA | 52 | 120~160 |
| SC11 | PP409560 | F: GACAATTATGATTGCGATTGGTT R: TTGATACCCTTCATTCCAGCTTA | 53 | 120~150 |
| SC13 | PP409560 | F: ACAACGCCAGCAGTAACCTATAA R: CGGTTGTGGAAGCAGTAGTAGTAG | 54 | 100~140 |
| SC17 | PP409562 | F: CACCTCTGTCTGAAAAACAAAAA R: AGGGACACCCAGTACACATCTAT | 50 | 130~170 |
| SC19 | PP409563 | F: CAAAACCGGTTGGTAATGAATAA R: TTTTGTTGTTGTTTTGTTGTTGC | 55 | 120~160 |
表3 9个缢蛏群体的平均遗传多样参数Tab. 3 Average genetic diversity index of 9 populations of S. constricta |
| 群体 | 参数 | SC03 | SC04 | SC05 | SC06 | SC07 | SC08 | SC11 | SC13 | SC17 | SC19 | 均值 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DD | Na | 3.0000 | 3.0000 | 4.0000 | 5.0000 | 6.0000 | 5.0000 | 4.0000 | 4.0000 | 4.0000 | 4.0000 | 4.2000 |
| Ne | 1.7527 | 1.9674 | 2.3810 | 4.3239 | 2.9268 | 3.4118 | 3.2200 | 1.8127 | 1.4207 | 2.2161 | 2.5433 | |
| Ho | 0.6000 | 0.0357 | 0.1000 | 0.4138 | 0.3333 | 0.6207 | 0.7667 | 0.2667 | 0.2333 | 0.0690 | 0.3439 | |
| He | 0.4367 | 0.5006 | 0.5898 | 0.7822 | 0.6695 | 0.7193 | 0.7011 | 0.4559 | 0.3011 | 0.5584 | 0.5715 | |
| PIC | 0.4510 | 0.4130 | 0.7330 | 0.7730 | 0.6070 | 0.9080 | 0.6650 | 0.4390 | 0.2870 | 0.4580 | 0.5734 | |
| P-value | 1.0000 | 0.0000* | 0.0000* | 0.0000* | 0.0033 | 0.0042 | 0.8245 | 0.0002* | 0.0561 | 0.0000* | 0.1888 | |
| QHD | Na | 5.0000 | 3.0000 | 5.0000 | 5.0000 | 4.0000 | 5.0000 | 3.0000 | 5.0000 | 4.0000 | 2.0000 | 4.1000 |
| Ne | 2.2416 | 1.4551 | 3.5644 | 4.4888 | 2.9851 | 3.3835 | 2.0619 | 1.6949 | 1.2721 | 1.9627 | 2.5110 | |
| Ho | 0.7333 | 0.0333 | 0.5333 | 0.3667 | 0.4667 | 0.6000 | 0.9667 | 0.4000 | 0.2333 | 0.0345 | 0.4368 | |
| He | 0.5633 | 0.3181 | 0.7316 | 0.7904 | 0.6763 | 0.7164 | 0.5237 | 0.4169 | 0.2175 | 0.4991 | 0.5453 | |
| PIC | 0.5690 | 0.2860 | 0.7040 | 0.7550 | 0.6230 | 0.8260 | 0.3980 | 0.4050 | 0.2050 | 0.3700 | 0.5141 | |
| P-value | 0.9964 | 0.0000* | 0.0000* | 0.0641 | 0.0025 | 1.0000 | 0.0459 | 1.0000 | 0.0000* | 0.0205 | 0.3129 | |
| ZH | Na | 4.0000 | 3.0000 | 6.0000 | 5.0000 | 4.0000 | 6.0000 | 3.0000 | 5.0000 | 3.0000 | 3.0000 | 4.2000 |
| Ne | 2.5388 | 1.1450 | 2.9561 | 4.1570 | 2.5678 | 4.5113 | 2.3866 | 1.9759 | 1.2685 | 1.4019 | 2.4909 | |
| Ho | 0.2667 | 0.0000 | 0.4138 | 0.3667 | 0.2000 | 0.6000 | 0.6071 | 0.4000 | 0.2333 | 0.0000 | 0.3088 | |
| He | 0.6164 | 0.1288 | 0.6733 | 0.7723 | 0.6209 | 0.7915 | 0.5916 | 0.5023 | 0.2153 | 0.2915 | 0.5204 | |
| PIC | 0.5360 | 0.1230 | 0.6750 | 0.7330 | 0.5300 | 0.8940 | 0.5080 | 0.4630 | 0.1990 | 0.2600 | 0.4921 | |
| P-value | 0.0205 | 0.0003* | 0.0022 | 0.0000* | 0.0001* | 0.0000* | 0.0386 | 0.2979 | 1.0000 | 0.0000* | 0.1359 | |
| QD | Na | 5.0000 | 2.0000 | 4.0000 | 5.0000 | 3.0000 | 4.0000 | 3.0000 | 5.0000 | 2.0000 | 4.0000 | 3.7000 |
| Ne | 1.6559 | 1.0689 | 3.4682 | 4.5421 | 1.3072 | 3.4221 | 2.0202 | 1.3206 | 1.0339 | 1.6408 | 2.1480 | |
| Ho | 0.4000 | 0.0000 | 0.5333 | 0.2963 | 0.2000 | 0.7000 | 0.8667 | 0.2000 | 0.0333 | 0.1333 | 0.3363 | |
| He | 0.4028 | 0.0655 | 0.7237 | 0.7945 | 0.239 | 0.7198 | 0.5136 | 0.2469 | 0.0333 | 0.3972 | 0.4136 | |
| PIC | 0.3770 | 0.0620 | 0.0730 | 0.7560 | 0.2220 | 0.8590 | 0.3930 | 0.2340 | 0.0320 | 0.3650 | 0.3373 | |
| P-value | 0.0316 | 0.0169 | 0.0036 | 0.0000* | 0.2409 | 0.0000* | 1.0000 | 0.0146 | 0.0000* | 0.0002* | 0.1308 | |
| LYG | Na | 1.0000 | 1.0000 | 5.0000 | 2.0000 | 2.0000 | 6.0000 | 4.0000 | 5.0000 | 3.0000 | 3.0000 | 3.2000 |
| Ne | 1.0000 | 1.0000 | 1.9956 | 1.2596 | 1.9978 | 4.2254 | 3.1634 | 1.7787 | 1.2237 | 1.1443 | 1.8789 | |
| Ho | 0.0000 | 0.0000 | 0.3667 | 0.2333 | 0.9667 | 0.7333 | 0.6333 | 0.3667 | 0.1333 | 0.0667 | 0.3500 | |
| He | 0.0000 | 0.0000 | 0.5073 | 0.2096 | 0.5079 | 0.7763 | 0.6955 | 0.4452 | 0.1859 | 0.1282 | 0.3456 | |
| PIC | 0.0000 | 0.0320 | 0.6650 | 0.1850 | 0.3750 | 0.8710 | 0.6190 | 0.4160 | 0.1800 | 0.1240 | 0.3467 | |
| P-value | 0.0000* | 1.0000 | 0.0000* | 1.0000 | 1.0000 | 0.1941 | 0.2597 | 0.0006* | 0.0669 | 0.0169 | 0.3538 | |
| NB | Na | 5.0000 | 2.0000 | 5.0000 | 5.0000 | 2.0000 | 5.0000 | 3.0000 | 5.0000 | 2.0000 | 4.0000 | 3.8000 |
| Ne | 3.1746 | 1.0689 | 3.3835 | 3.1142 | 1.4274 | 4.0909 | 2.2727 | 2.2814 | 1.0339 | 1.4718 | 2.3319 | |
| Ho | 0.4667 | 0.0000 | 0.4667 | 0.4333 | 0.3667 | 0.5333 | 0.9333 | 0.4000 | 0.0333 | 0.1000 | 0.3733 | |
| He | 0.6966 | 0.0655 | 0.7164 | 0.6904 | 0.3045 | 0.7684 | 0.5695 | 0.5712 | 0.0333 | 0.3260 | 0.4742 | |
| PIC | 0.6560 | 0.0620 | 0.8160 | 0.6430 | 0.2550 | 0.9030 | 0.4610 | 0.5500 | 0.0640 | 0.3050 | 0.4715 | |
| P-value | 0.1371 | 0.0169 | 0.0000* | 0.0058 | 1.0000 | 0.0217 | 0.9827 | 0.0000* | 1.0000 | 0.0000* | 0.3164 | |
| XM | Na | 2.0000 | 1.0000 | 4.0000 | 5.0000 | 5.0000 | 5.0000 | 3.0000 | 4.0000 | 4.0000 | 5.0000 | 3.8000 |
| Ne | 1.6000 | 1.0000 | 2.2876 | 3.5573 | 1.9063 | 3.8793 | 2.2032 | 2.1102 | 1.4634 | 1.8653 | 2.1873 | |
| Ho | 0.4333 | 0.0000 | 0.3462 | 0.3667 | 0.4333 | 0.8000 | 0.9333 | 0.4333 | 0.2000 | 0.1000 | 0.4046 | |
| He | 0.3814 | 0.0000 | 0.5739 | 0.7311 | 0.4836 | 0.7548 | 0.5554 | 0.5350 | 0.3220 | 0.4718 | 0.4809 | |
| PIC | 0.6720 | 0.4410 | 0.8840 | 0.4420 | 0.4410 | 0.8840 | 0.4420 | 0.5260 | 0.2950 | 0.4440 | 0.5471 | |
| P-value | 0.0000* | 0.0044 | 0.6226 | 0.8616 | 0.0044 | 0.6226 | 0.8616 | 0.0068 | 0.0048 | 0.0000* | 0.2989 | |
| HZ | Na | 5.0000 | 3.0000 | 5.0000 | 5.0000 | 2.0000 | 5.0000 | 4.0000 | 4.0000 | 2.0000 | 4.0000 | 3.9000 |
| Ne | 2.2939 | 1.2649 | 2.4931 | 3.4417 | 1.5139 | 3.8710 | 2.8257 | 1.3657 | 1.0339 | 1.5139 | 2.1618 | |
| Ho | 0.5333 | 0.0333 | 0.2333 | 0.5667 | 0.4333 | 0.6333 | 0.7667 | 0.3000 | 0.0333 | 0.1333 | 0.3667 | |
| He | 0.5734 | 0.2130 | 0.6090 | 0.7215 | 0.3452 | 0.7542 | 0.6571 | 0.2723 | 0.0333 | 0.3452 | 0.4524 | |
| PIC | 0.5880 | 0.2130 | 0.7830 | 0.7260 | 0.3520 | 0.9010 | 0.6570 | 0.2450 | 0.0330 | 0.3460 | 0.4844 | |
| P-value | 0.0937 | 0.0001* | 0.0000* | 0.0683 | 1.0000 | 0.0000* | 0.6531 | 1.0000 | 0.0000* | 0.0263 | 0.2841 | |
| BH | Na | 5.0000 | 3.0000 | 5.0000 | 3.0000 | 5.0000 | 5.0000 | 5.0000 | 5.0000 | 3.0000 | 4.0000 | 4.3000 |
| Ne | 2.2032 | 1.2270 | 2.4658 | 1.3943 | 4.5226 | 1.8927 | 2.8939 | 2.8571 | 1.9231 | 1.7561 | 2.3136 | |
| Ho | 0.6333 | 0.2000 | 0.4333 | 0.3333 | 0.7667 | 0.5333 | 0.6667 | 0.7667 | 0.6667 | 0.5333 | 0.5533 | |
| He | 0.5554 | 0.1881 | 0.6045 | 0.2876 | 0.7921 | 0.4797 | 0.6655 | 0.6610 | 0.4881 | 0.4379 | 0.5160 | |
| PIC | 0.5030 | 0.1770 | 0.7760 | 0.2510 | 0.8590 | 0.6010 | 0.7320 | 0.7160 | 0.6220 | 0.7520 | 0.5989 | |
| P-value | 0.9440 | 1.0000 | 0.0181 | 1.0000 | 0.4223 | 0.7689 | 0.9183 | 1.0000 | 0.9940 | 0.9761 | 0.8042 |
注: 等位基因总数为352个。*表示经过Bonferroni矫正后显著偏离哈迪-温伯格定律(P < 0.01) |
表4 9个缢蛏群体间的遗传分化系数(对角线下)和基因流(对角线以上)Tab. 4 Genetic differentiation index (below diagonal) and gene flow (above diagonal) of 9 populations of S. constricta |
| 群体 | DD | QHD | ZH | QD | LYG | NB | XM | HZ | BH |
|---|---|---|---|---|---|---|---|---|---|
| DD | — | 2.7297 | 1.6497 | 1.0460 | 0.5298 | 0.8741 | 0.8756 | 0.8188 | 0.8586 |
| QHD | 0.0839 | — | 4.3204 | 1.5281 | 0.6187 | 1.1282 | 1.4507 | 0.9450 | 0.7350 |
| ZH | 0.1316 | 0.0547 | — | 2.6981 | 0.7194 | 1.5093 | 1.2707 | 0.8271 | 0.8508 |
| QD | 0.1929 | 0.1406 | 0.0848 | — | 0.7186 | 1.2951 | 0.9248 | 1.0665 | 0.5406 |
| LYG | 0.3206 | 0.2878 | 0.2579 | 0.2581 | — | 1.7892 | 1.0146 | 1.0294 | 0.4620 |
| NB | 0.2224 | 0.1814 | 0.1421 | 0.1618 | 0.1226 | — | 2.1219 | 1.1939 | 0.6296 |
| XM | 0.2221 | 0.1470 | 0.1644 | 0.2128 | 0.1977 | 0.1054 | — | 3.4211 | 0.6309 |
| HZ | 0.2339 | 0.2092 | 0.2321 | 0.1899 | 0.1954 | 0.1142 | 0.0681 | — | 0.6009 |
| BH | 0.2255 | 0.2538 | 0.2721 | 0.3162 | 0.3511 | 0.2842 | 0.2838 | 0.2938 | — |
表5 9个缢蛏群体的AMOVA分析Tab. 5 AMOVA analysis of 9 populations of S. constricta |
| 变异来源 | 自由度 | 平方和 | 方差分量 | 变异百分比/% |
|---|---|---|---|---|
| 群体间 | 8 | 630.102 | 1.26981Va | 33.04** |
| 群体内 | 531 | 1366.717 | 2.57385Vb | 66.96** |
| 总计 | 539 | 1996.819 | 3.84367 | 100 |
注: **表示1023次模拟检验后显示为差异极显著(p < 0.01)。Va表示 群体间的遗传变异, Vb表示群体内的遗传变异 |
表6 9个群体间的Nei’s遗传距离(对角线下)和遗传相似指数(对角线上)Tab. 6 Nei’s genetic distance (below diagonal) and genetic identity (above diagonal) of 9 populations |
| 群体 | DD | QHD | ZH | QD | LYG | NB | XM | HZ | BH |
|---|---|---|---|---|---|---|---|---|---|
| DD | — | 0.7501 | 0.6262 | 0.5149 | 0.1806 | 0.3759 | 0.3309 | 0.3403 | 0.1000 |
| QHD | 0.2875 | — | 0.8632 | 0.6573 | 0.2957 | 0.5220 | 0.5833 | 0.4370 | 0.0712 |
| ZH | 0.4681 | 0.1471 | — | 0.8187 | 0.4218 | 0.6405 | 0.5575 | 0.4148 | 0.0680 |
| QD | 0.6637 | 0.4196 | 0.2000 | — | 0.4845 | 0.6434 | 0.4846 | 0.5954 | 0.0613 |
| LYG | 1.7115 | 1.2185 | 0.8631 | 0.7247 | — | 0.7722 | 0.5684 | 0.5911 | 0.0559 |
| NB | 0.9785 | 0.6501 | 0.4455 | 0.4411 | 0.2586 | — | 0.7310 | 0.7133 | 0.0792 |
| XM | 1.1059 | 0.5391 | 0.5843 | 0.7244 | 0.5649 | 0.3134 | — | 0.8574 | 0.0659 |
| HZ | 1.0780 | 0.8278 | 0.8798 | 0.5185 | 0.5257 | 0.3378 | 0.1538 | — | 0.0566 |
| BH | 2.3022 | 2.6421 | 2.6885 | 2.7917 | 2.8838 | 2.5353 | 2.7198 | 2.8723 | — |
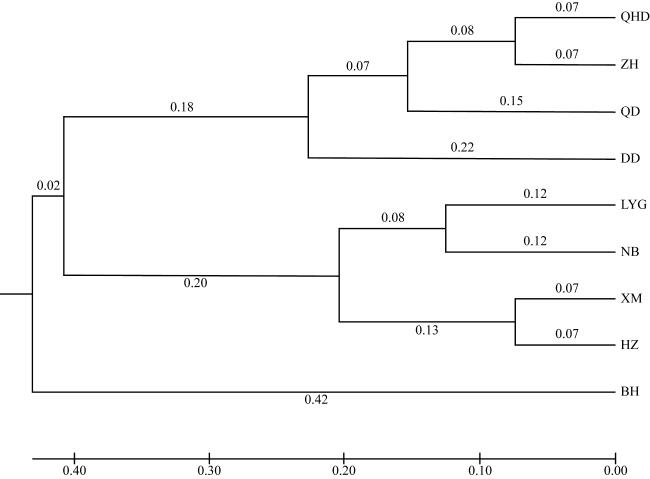
图2 基于缢蛏9个群体遗传距离构建的UPGMA聚类图进化树上的数值及长度表示不同地理群体缢蛏之间的进化距离 Fig. 2 Construction of UPMGA Cluster Map based on genetic distance of 9 populations of S. constricta The values and lengths on the evolutionary tree represent the evolutionary distance between different geographic populations of razor clams |
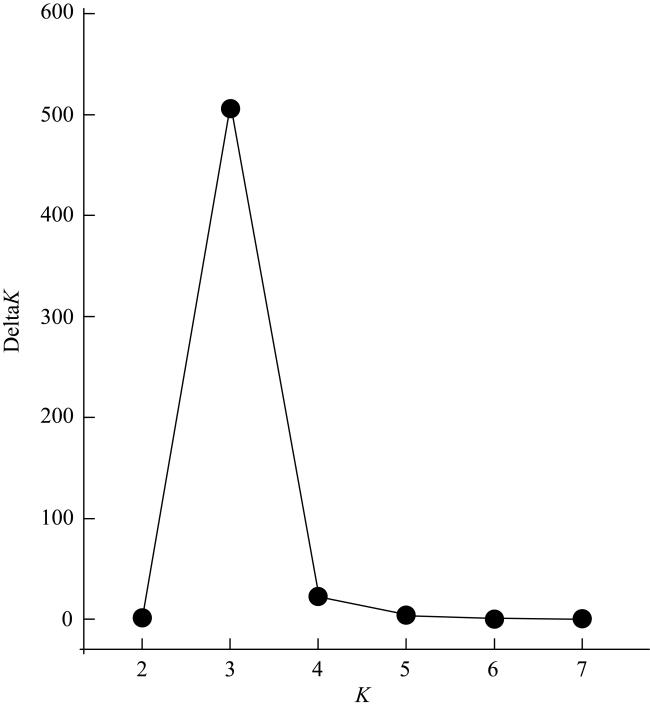
图4 根据Structure Harvester评估的最佳K值Fig. 4 The optimal K value determined by Structure Harvester evaluation |
| [1] |
包文斌, 束婧婷, 许盛海, 等, 2007. 样本量和性比对微卫星分析中群体遗传多样性指标的影响[J]. 中国畜牧杂志, 43(1): 6-9.
|
| [2] |
范士琦, 冯婧昀, 苗晓敏, 等, 2023. 重庆养殖场鳜群体微卫星遗传多样性研究[J]. 水产养殖, 44(7): 18-23.
|
| [3] |
谷德贤, 王婷, 许玉甫, 等, 2021. 利用微卫星分子标记分析渤海湾的口虾蛄遗传多样性[J]. 水产科学, 40(5): 693-699.
|
| [4] |
郭香, 曾志南, 郑雅友, 等, 2018. 福建牡蛎选育群体的遗传多样性[J]. 中国水产科学, 25(5): 1131-1136.
|
| [5] |
黄小林, 李文俊, 林黑着, 等, 2018. 基于线粒体DNA D-loop序列的黄斑篮子鱼群体遗传多样性分析[J]. 热带海洋学报, 37(4): 45-51.
|
| [6] |
黄新芯, 刘玉萍, 宁子君, 等, 2025. 基于微卫星标记的中国近海龙头鱼群体遗传结构分析[J]. 水生态学杂志, 46(1): 90-98.
|
| [7] |
李成华, 2004. 泥蚶分子系统分化及缢蛏遗传多样性的研究[D]. 青岛: 中国科学院研究生院(海洋研究所): 53-62 (in Chinese).
|
| [8] |
李景芬, 夏正龙, 栾生, 等, 2020. 五个罗氏沼虾群体遗传多样性的微卫星分析[J]. 水生生物学报, 44(6): 1208-1214.
|
| [9] |
李鸥, 赵莹莹, 郭娜, 等, 2009. 草鱼种群SSR分析中样本量及标记数量对遗传多度的影响[J]. 动物学研究, 30(2): 121-130.
|
| [10] |
李妍, 姚健涛, 张恩烁, 等, 2024. 长牡蛎(Crassostrea gigas)野生与选育群体的微卫星遗传多样性分析[J]. 海洋与湖沼, 55(2): 462-470.
|
| [11] |
李镒民, 丁琳琳, 谭杰, 等, 2019. 马氏珠母贝F8代黑壳色选育系群体与普通养殖群体遗传多样性分析[J]. 基因组学与应用生物学, 38(11): 4961-4967.
|
| [12] |
梁园, 付敬强, 沈铭辉, 等, 2022. 方斑东风螺3个选育世代遗传多样性和遗传结构的微卫星分析[J]. 海洋科学, 46(10): 85-93.
|
| [13] |
刘博, 邵艳卿, 王侃, 等, 2013. 4个缢蛏群体遗传多样性和系统发生关系的微卫星分析[J]. 海洋科学, 37(8): 96-102.
|
| [14] |
刘达博, 牛东红, 冯冰冰, 等, 2011. 乐清湾和三沙湾缢蛏群体遗传多样性的微卫星分析[J]. 上海海洋大学学报, 20(3): 350-357.
|
| [15] |
牛东红, 李家乐, 冯冰冰, 等, 2009. 缢蛏6个群体遗传结构的ISSR分析[J]. 应用与环境生物学报, 15(3): 332-336.
|
| [16] |
牛东红, 陈慧, 林国文, 等, 2010. 缢蛏群体微卫星分析中样本量对遗传多样性指标的影响[J]. 海洋科学进展, 28(2): 203-208.
|
| [17] |
牛东红, 冯冰冰, 刘达博, 等, 2011. 浙闽沿海缢蛏群体遗传结构的微卫星和线粒体CO Ⅰ序列分析[J]. 水产学报, 35(12): 1805-1813.
|
| [18] |
彭敏, 肖珊, 洪传远, 等, 2022. 华南沿海长鳍篮子鱼不同地理群体的遗传多样性分析[J]. 水生态学杂志, 43(5): 127-133.
|
| [19] |
阮惠婷, 徐姗楠, 李敏, 等, 2020. 飘鱼微卫星位点的筛选及珠江流域5个地理群体的遗传多样性分析[J]. 水生生物学报, 44(3): 501-508.
|
| [20] |
邵艳卿, 方军, 柏艳, 等, 2015. 缢蛏(Sinonovacula constricta) EST-SSR标记与生长性状的相关性分析[J]. 海洋与湖沼, 46(5): 1146-1152.
|
| [21] |
苏晓盈, 代永仙, 谭杰, 等, 2021. 马氏珠母贝(Pinctada martensii)黑壳色养殖群体SSR遗传多样性分析[J]. 基因组学与应用生物学, 40(2): 615-621.
|
| [22] |
孙志鹏, 鲁翠云, 那荣滨, 等, 2023. 利用线粒体序列比较分析梭鲈鸭绿江和乌伦古湖群体的遗传结构[J]. 水产学杂志, 36(5): 10-16, 26.
|
| [23] |
唐芳, 温贝妮, 刘红, 2021. 不同凡纳滨对虾养殖群体的微卫星遗传多样性分析[J]. 南方农业学报, 52(4): 1108-1115.
|
| [24] |
滕爽爽, 胡高宇, 范建勋, 等, 2021. 缢蛏5个群体遗传多样性和遗传分化的SNP分析[J]. 水生生物学报, 45(4): 861-870.
|
| [25] |
田镇, 陈爱华, 曹奕, 等, 2021. 红壳色文蛤选育群体遗传多样性的微卫星分析[J]. 南方农业学报, 52(9): 2582-2589.
|
| [26] |
王冬群, 李太武, 苏秀榕, 2005. 象山缢蛏养殖群体和野生群体遗传多样性的比较[J]. 中国水产科学, 12(2): 138-143.
|
| [27] |
吴玲, 2013. 马氏珠母贝和缢蛏的遗传多样性研究[D]. 青岛: 中国海洋大学: 49-86.
|
| [28] |
徐义平, 许会宾, 金凯, 等, 2017. 浙江乐清湾缢蛏的形态和遗传多样性[J]. 上海海洋大学学报, 26(1): 31-37.
|
| [29] |
于思梦, 高磊, 王伟, 等, 2017. 辽宁黄渤海沿岸长牡蛎遗传多样性分析[J]. 经济动物学报, 21(4): 215-220.
|
| [30] |
于颖, 孟祥盈, 王秀利, 等, 2007. 缢蛏遗传多样性的RAPD分析[J]. 生物技术通报, (6): 138-140.
|
| [31] |
张帝, 强俊, 傅建军, 等, 2022. 基于微卫星标记和线粒体D-loop序列的5个大口黑鲈群体遗传变异分析[J]. 中国水产科学, 29(9): 1277-1289.
|
| [32] |
张秀英, 张晓军, 赵翠, 等, 2012. 栉孔扇贝BES-SSR的开发及遗传多样性分析[J]. 水产学报, 36(6): 815-824.
|
| [33] |
赵文浩, 易少奎, 周琼, 等, 2023. 新疆塔里木河叶尔羌高原鳅群体遗传学研究[J]. 水产科学, 42(4): 664-673.
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}