
珊瑚共附生嗜盐单胞菌原噬菌体上的HicAB毒素-抗毒素系统的鉴定及其结构分析
|
张雨(1999—), 男, 江西省上饶市人, 硕士, 从事海洋微生物生理与生化研究。email: yzhang0517@126.com |
Copy editor: 林强
收稿日期: 2025-02-17
修回日期: 2025-03-14
网络出版日期: 2025-03-18
基金资助
国家自然科学基金项目(42188102)
中国科学院南海海洋研究所专项基金(SCSIO2023QY03)
海洋负排放国际大科学计划(ONCE)
海南大学南海海洋资源利用国家重点实验室开放项目(MRUKF2023001)
Identification and structural analysis of the HicAB toxin-antitoxin system encoded by a prophage in coral-associated Halomonas meridiana
Received date: 2025-02-17
Revised date: 2025-03-14
Online published: 2025-03-18
Supported by
National Natural Science Foundation of China(42188102)
Special Fund of the South China Sea Institute of Oceanology, Chinese Academy of Sciences(SCSIO2023QY03)
Ocean Negative Carbon Emissions Program(ONCE)
Open project of State Key Laboratory of South China Sea Marine Resources Utilization, Hainan University(MRUKF2023001)
噬菌体与细菌宿主的相互作用对珊瑚健康和珊瑚礁稳定性具有重要影响, 也是病毒学领域的研究热点之一。为应对噬菌体的感染压力, 细菌进化出多种固有和适应性免疫系统, 其中毒素-抗毒素(toxin-antitoxin, TA)系统是重要的防御机制之一。本研究通过生物信息学分析预测, 珊瑚共附生嗜盐单胞菌Halomonas meridiana SCSIO 43005 (Hm43005)所携带的原噬菌体Phm2中的CTT34_05265和CTT34_05260基因分别编码HmHicA和HmHicB蛋白, 并通过大肠杆菌生长实验证实其构成TA系统。Pull-down和细菌双杂交实验进一步验证了HmHicA与HmHicB之间的相互作用。凝胶迁移阻滞实验(electrophoretic mobility shift assay, EMSA)和DNaseⅠ足迹实验表明, HmHicB通过特异性结合hicAB启动子-35区上游和-10区的回文序列, 发挥转录自调控功能。此外, HmHicB单独表达对宿主表现出轻微毒性, 提示抗毒素蛋白在细胞内可能具有其他调控靶点。结构分析显示, HmHicA具有核糖体非依赖性RNA切割酶活性, 而HmHicB包含N端毒素结合结构域和C端DNA结合结构域。HmHicB通过C端形成同源二聚体, 并与HmHicA以2:2的化学计量比形成复合物。HmHicA带正电荷的活性口袋与HmHicB带负电荷的毒素结合结构域之间的相互作用, 以及HmHicA His24活性位点的包埋, 可能是抗毒素抑制毒素活性的分子机制。这些研究结果为深入解析该TA系统的性质及其生理生态功能奠定了重要基础。
张雨 , 刘自尧 , 王晓雪 , 陈冉 . 珊瑚共附生嗜盐单胞菌原噬菌体上的HicAB毒素-抗毒素系统的鉴定及其结构分析[J]. 热带海洋学报, 2025 , 44(6) : 120 -131 . DOI: 10.11978/2025022
The interaction between phages and bacterial hosts significantly impacts coral health and reef stability, representing a key focus in virology. To combat phage infection, bacteria have evolved diverse innate and adaptive immune systems, including toxin-antitoxin (TA) systems, a crucial defense mechanism. In this study, bioinformatics analysis predicted that the CTT34_05265 and CTT34_05260 genes within the prophage Phm2 of the coral-associated bacterium Halomonas meridiana SCSIO 43005 (Hm43005) encode HmHicA and HmHicB proteins, respectively. Their TA system functionality was confirmed through E. coli growth assays. Pull-down and bacterial two-hybrid experiments validated the interaction between HmHicA and HmHicB. Electrophoretic mobility shift assays (EMSA) and DNase I footprinting revealed that HmHicB specifically binds to palindromic sequences upstream of the -35 and -10 regions of the hicAB promoter, mediating transcriptional autoregulation. Additionally, HmHicB alone exhibited mild toxicity, suggesting potential alternative regulatory targets for the antitoxin. Structural analysis indicated that HmHicA functions as a ribosome-independent RNase, while HmHicB contains an N-terminal toxin-binding domain and a C-terminal DNA-binding domain. HmHicB forms a homodimer via its C-terminus and assembles with HmHicA in a 2:2 stoichiometric complex. The molecular mechanism of antitoxin-mediated toxin inhibition likely involves electrostatic interactions between the positively charged active pocket of HmHicA and the negatively charged toxin-binding domain of HmHicB, as well as the burial of the HmHicA His24 active site. These findings provide a foundation for further exploration of the properties and physiological roles of this TA system.
表1 本文所用菌株Tab. 1 Bacterial strains used in this study |
| 菌株 | 描述 | 来源 |
|---|---|---|
| Hm43005 | Isolated from Coral samples of G. fascicularis. | 实验室保存 |
| WM3064 | thrB1004 pro thi rpsL hsdS lacZΔM15 RP4-360 Δ(araBAD)567 ΔdapA1341:: [erm pir] | 实验室保存 |
| BL21 (DE3) | F-ompT hsdSB(rB-mB-) gal dcm λ(DE3) Ω PtacUV5::T7 polymerase | Novagen |
| BW25113 (K-12) | lacIq rrnBT14 ΔlacZWJ16 hsdR514 ΔaraBADAH33 ΔrhaBADLD78 rph-1 | Baba et al, 2006 |
| Top10 | F-, mcrAΔ(mrr-hsd RMS-mcrBC), ϕ80, lacZΔM15, ΔlacX74, recA1, araΔ139Δ(ara-leu)7697, galU, galK, rps, (Strr) endA1, nupG | 实验室保存 |
注: KmR, CmR, AmpR分别代表卡那霉素、氯霉素和氨苄青霉素。 |
表2 本文所用质粒Tab. 2 Plasmids used in this study |
| 质粒 | 描述 | 来源 |
|---|---|---|
| pHGECm | CmR, IPTG inducible expression vector in E.coli and H.meridiana SCSIO 43005 strains | 实验室保存 |
| pHGECm-hicA | CmR; over-expression vector for CTT34_05265 | 本研究 |
| pHGECm-hicB | CmR; over-expression vector for CTT34_05260 | 本研究 |
| pHGECm-hicAB | CmR; over-expression vector for CTT34_05260 and CTT34_05265 as one operon | 本研究 |
| pUT18C-hicA | AmpR; interaction assay vector for CTT34_05265 | 本研究 |
| pKT25-hicB | KmR; interaction assay vector for CTT34_05260 | 本研究 |
| pET28b | KmR, IPTG inducible expression and purified vector | Novagen |
| pET28b-hicA-his | KmR; pET28b PT7-lac::hicA with C-terminal His-tagged | 本研究 |
| pET28b-hicB-his | KmR; pET28b PT7-lac::hicB with C-terminal His-tagged | 本研究 |
| pET28b-his-hicAB | KmR; pET28b PT7-lac:: hicAB with N-terminal His-tagged | 本研究 |
注: KmR, CmR, AmpR分别代表卡那霉素、氯霉素和氨苄青霉素。 |
表3 引物信息Tab. 3 Sequences of designed primers used in this study |
| 引物名称 | 序列(5′-3′) | 用途 |
|---|---|---|
| pHGECm-hicA-F | TAACAATTTCACACAGGAGAGATGAACAGCAGAGCACTGATC | 毒性验证 |
| pHGECm-hicA-R | ATCCGCCAAAACAGCCAAGCTTCATTCGAGGCCAGCGCTTTTC | 毒性验证 |
| pHGECm-hicB-F | TAACAATTTCACACAGGAGAGATGTTATTTCCCATTGCCATTG | 毒性验证 |
| pHGECm-hicB-R | ATCCGCCAAAACAGCCAAGCTTTAGCTGGCTTGTTTGTTCCTG | 毒性验证 |
| pUT18C-hicA-F | ACTCTAGAGGATCCCCGGGTACCGATGAACAGCAGAGCACTGATC | 细菌双杂交 |
| pUT18C-hicA-R | ATTACTTAGTTATATCGATGAATTTCATTCGAGGCCAGCGCTTTTC | 细菌双杂交 |
| pKT25-hicB-F | CTAGAGGATCCCCGGGTACCTATGTTATTTCCCATTGCCATTG | 细菌双杂交 |
| pKT25-hicB-R | GAATTCTTAGTTACTTAGTTAGCTGGCTTGTTTGTTCCTG | 细菌双杂交 |
| pET28b-hicA-his-F | CTTTAAGAAGGAGATATACCATGAACAGCAGAGCACTGATC | 蛋白表达纯化 |
| pET28b-hicA-his-R | CTCGAGTGCGGCCGCAAGCTTTTCGAGGCCAGCGCTTTTCC | 蛋白表达纯化 |
| pET28b-hicB-his-F | CTTTAAGAAGGAGATATACCATGTTATTTCCCATTGCCATTG | 蛋白表达纯化 |
| pET28b-hicB-his-R | CTCGAGTGCGGCCGCAAGCTTGCTGGCTTGTTTGTTCCTGGC | 蛋白表达纯化 |
| pET28b-his-hicAB-F | GGTGGACAGCAAATGGGTCGGATGAACAGCAGAGCACTGATC | Pull-down分析 |
| pET28b-his-hicAB-R | CTCGAGTGCGGCCGCAAGCTTTTAGCTGGCTTGTTTGTTCCTG | Pull-down分析 |
注: “F”表示上游引物, “R”表示下游引物。 |
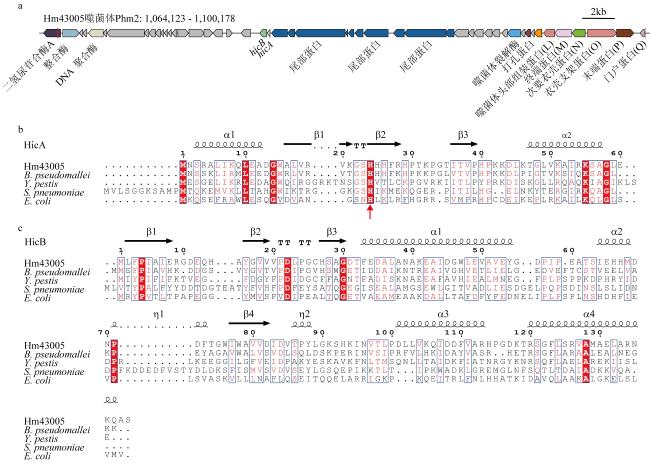
图1 HicAB系统的生物信息学分析a. hicAB基因在原噬菌体中的位置, hicA和hicB分别采用蓝色和绿色显示; b. Hm43005菌株编码的HmHicA与已解析结构的B. pseudomallei (PDB 6G26)、Y. pestis (PDB 4P78)、S. pneumoniae (PDB 5YRZ)和E. coli (PDB 6HPB)四个物种HicA序列比对, c. Hm43005菌株编码的HmHicB与已解析结构的B. pseudomallei、Y. pestis、E. coli和S. pneumoniae四个物种HicB序列比对 Fig. 1 Bioinformatics analysis of the HicAB system. (a) The location of hicAB operon within the prophage, with hicA and hicB shown in blue and green, respectively; (b) sequence alignment of HmHicA encoded by the Hm43005 strain with the structurally resolved HicA sequences from B. pseudomallei, Y. pestis, S. pneumoniae, and E. coli; (c) sequence alignment of HmHicB encoded by the Hm43005 strain with the structurally resolved HicB sequences from B. pseudomallei, Y. pestis, E. coli and S. pneumoniae |
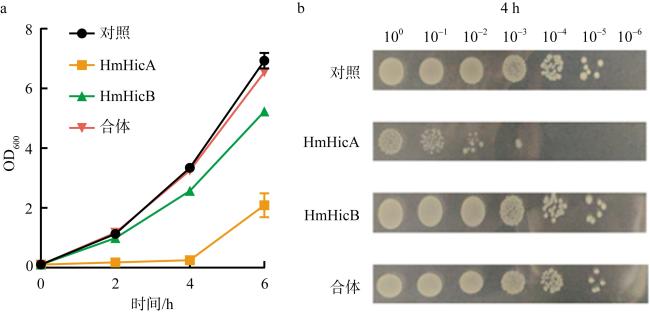
图2 大肠杆菌生长实验确认Phm2上的HmHicAB构成一对TA系统a. 生长曲线表明过表达毒素HmHicA可以抑制大肠杆菌生长, HmHicB可中和HmHicA的毒性; b. 存活实验(CFU·mL-1)证实了hicA和hicB组成TA系统 Fig. 2 Growth experiments of E. coli confirm that hicA and hicB on Phm2 constitute a TA system. (a) Growth curves indicate that overexpression of toxins can inhibit the growth of E. coli; (b) survival assays (CFU·mL-1) confirmed that hicA and hicB constitute a TA system |
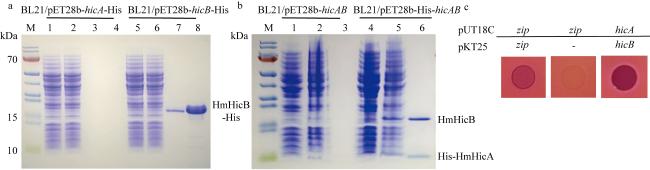
图3 实验验证HmHicA与HmHicB相互作用a. SDS-PAGE检测纯化到抗毒素HmHicB蛋白大小和纯度; 泳道1和5: 诱导前; 泳道2和6: 诱导后; 泳道3和7: 洗脱样1; 泳道4和8: 洗脱样2。b. SDS-PAGE显示表达毒素HmHicA蛋白可以成功下拉HmHicB抗毒素蛋白; 泳道1和4: 诱导前; 泳道2和5: 诱导后; 泳道3和6: 洗脱样1。c. 细菌双杂交鉴定HmHicA与HmHicB相互作用 Fig. 3 Experimental validation of the interaction between HmHicA and HmHicB. (a) SDS-PAGE analysis of the size and purity of the purified anti-toxin HmHicB protein: Lane 1 and 5 (pre-induction), Lane 2 and 6 (post-induction), Lane 3 and 7 (elution sample 1), Lane 4 and 8 (elution sample 2); (b) SDS-PAGE analysis demonstrates that the expressed toxin HicA protein successfully pulls down the HmHicB anti-toxin protein: Lane 1 and 4 (pre-induction), Lane 2 and 5 (post-induction), Lane 3 and 6 (elution sample 1); (c) bacterial two-hybrid assay confirms the interaction between HmHicA and HmHicB |
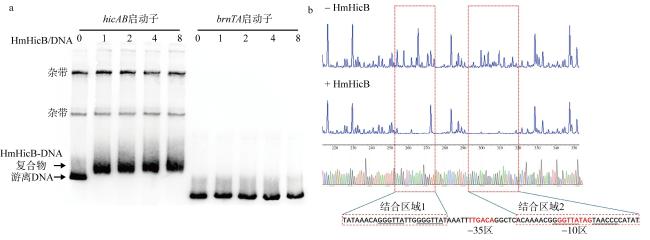
图4 体外纯化的HmHicB抗毒素蛋白可以与hicAB的启动子结合a. 抗毒素HmHicB能够结合自身启动子, 并且随着蛋白浓度增加结合能力增强, 0、1、2、4和8分别代表蛋白与核酸摩尔比(HmHicB/DNA) 为0∶1、1∶1、2∶1、4∶1、8∶1, 对照为Hm43005菌株brnTA系统的启动子; b. HmHicB与hicAB启动子-35区上游和-10区的回文序列结合, 对照为不加HmHicB的测序结果。-35和-10区序列用红色字体显示, 结合区域1和2用两个红色虚线方框显示, 重复和回文序列用下划线标记 Fig. 4 The in vitro purified HmHicB antitoxin protein can bind to the hicAB promoter. (a) The antitoxin HmHicB can bind to its own promoter, and its binding ability increases with higher protein concentrations. The numbers 0, 1, 2, 4, and 8 represent the molar ratios of protein to nucleic acid (HmHicB/DNA) at 0∶1, 1∶1, 2∶1, 4∶1, and 8∶1, respectively. The control is the promoter of the brnTA system from strain Hm43005. (b) HmHicB binds to the palindromic sequences upstream of the -35 and within the -10 regions of the hicAB promoter. The control is the sequencing result without HmHicB. The -35 and -10 regions are highlighted in red, and the binding regions 1 and 2 are marked with two red dashed boxes. Repeated and palindromic sequences are underlined |
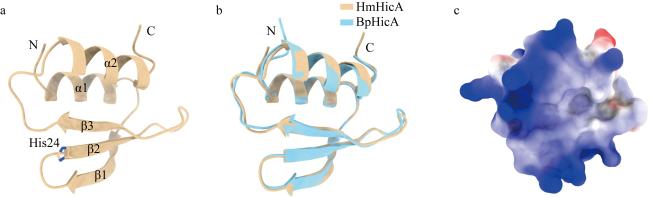
图5 HmHicA的结构特征分析a. AlphaFold 3预测的HmHicA单体结构; b. HmHicA与B. pseudomallei HicA结构对比分析; c. HmHicA表面电荷分析 Fig. 5 Structural analysis of HmHicA. (a) Monomer structure of HmHicA predicted by AlphaFold 3; (b) comparative structural analysis of HmHicA and B. pseudomallei HicA; (c) surface charge analysis of HmHicA |
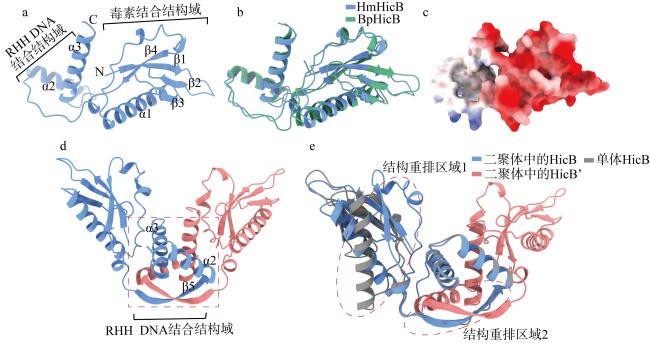
图6 HmHicB的C端相互作用形成典型的RHH DNA结合结构域a. AlphaFold 3预测的HmHicB单体结构; b. HmHicB与B. pseudomallei HicB结构对比分析; c. HmHicB表面电荷分析; d. AlphaFold 3预测的HmHicB二聚体结构, 二聚化形成的RHH结构域采用红色虚线方框标出; e. HmHicB单体与二聚体结构对比分析, 两处结构重排的区域采用红色虚线椭圆标出 Fig. 6 C-terminal interactions of HmHicB form a typical RHH DNA-binding domain. (a) Monomer structure of HmHicB predicted by AlphaFold 3; (b) comparative structural analysis of HmHicB and B. pseudomallei HicB; (c) surface charge analysis of HmHicB; (d) dimer structure of HmHicB predicted by AlphaFold 3, with the RHH domain formed by dimerization marked by a red dashed box; (e) comparative structural analysis of HmHicB monomer and dimer, with regions undergoing structural rearrangement marked by red dashed ellipses |
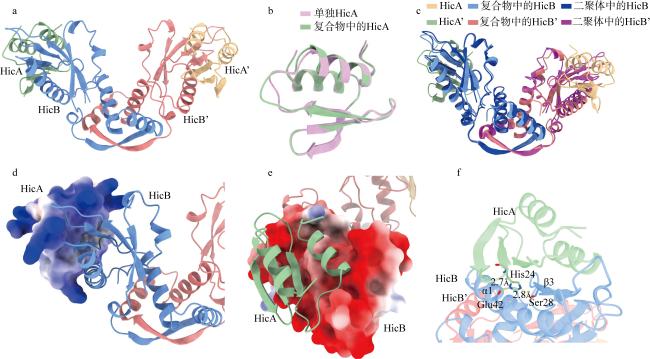
图7 HmHicAB复合物结构特征分析a. AlphaFold 3预测的HmHicA:HmHicB比例为2:2的复合物结构; b. HmHicA单体结构与TA复合物中的结构对比分析; c. HmHicB二聚体结构与TA复合物中的结构对比分析; d. TA复合物中HmHicA的表面电荷分析; e. TA复合物中HmHicB的表面电荷分析; f. TA复合物中HmHicA的关键活性位点His24所处周围环境分析 Fig. 7 Structural characterization of the HmHicAB complex. (a) Predicted 2:2 complex structure of HmHicA:HmHicB by AlphaFold 3; (b) structural comparison between the HmHicA monomer and its conformation in the TA complex; (c) structural comparison between the HmHicB dimer and its conformation in the TA complex; (d) surface charge analysis of HmHicA in the TA complex; (e) surface charge analysis of HmHicB in the TA complex; (f) analysis of the surrounding environment of the key active site His24 in HmHicA within the TA complex |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}