
升温对丛生盔形珊瑚两种形态型代谢和共生藻光合生理的影响
|
何茜(1997—), 女, 广东省茂名市人, 硕士研究生, 从事珊瑚生物学和珊瑚礁生态学研究。email: |
Copy editor: 殷波
收稿日期: 2021-06-22
修回日期: 2021-08-26
网络出版日期: 2021-08-27
基金资助
国家自然科学基金(41976120)
海南省重点研发计划(ZDYF2020200)
三亚崖州湾科技局科研项目(SKJC-2020-01-010)
广东省科技计划(2020B1212060058)
Effects of warming on the metabolism and photosynthetic physiology of the symbiotic algae of two morphological types of Galaxea fascicularis
Copy editor: YIN Bo
Received date: 2021-06-22
Revised date: 2021-08-26
Online published: 2021-08-27
Supported by
National Natural Science Foundation of China(41976120)
Key Research and Development Project of Hainan Province, China(ZDYF2020200)
Research Project of Sanya Yazhou-Bay SCI-TECH City Administration(SKJC-2020-01-010)
Science and Technology Planning Project of Guangdong Province, China(2020B1212060058)
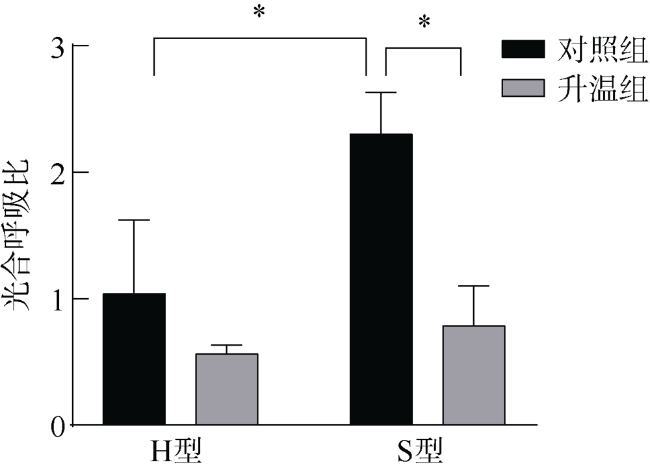
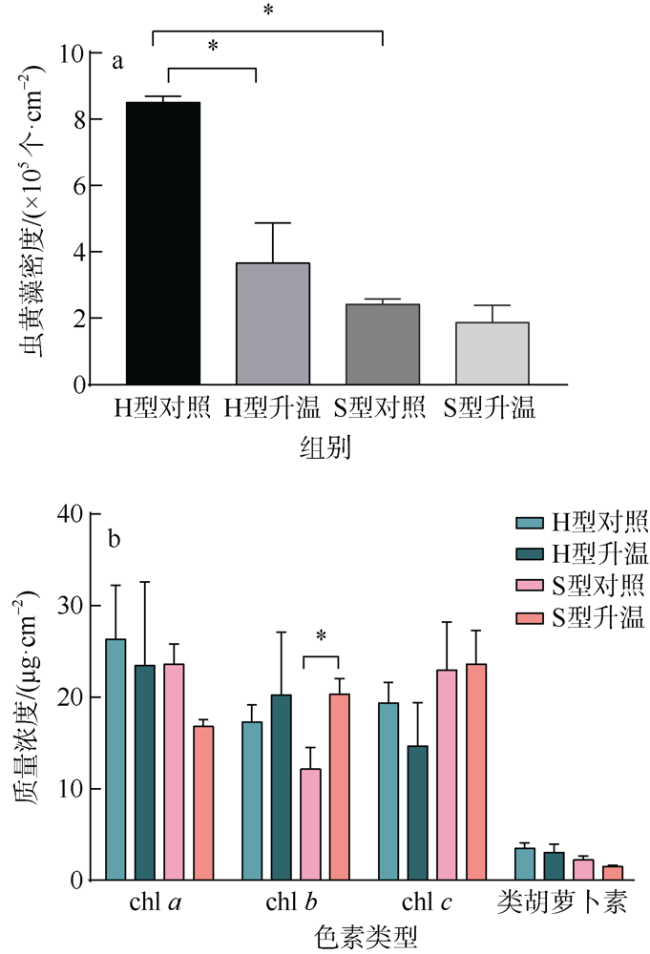
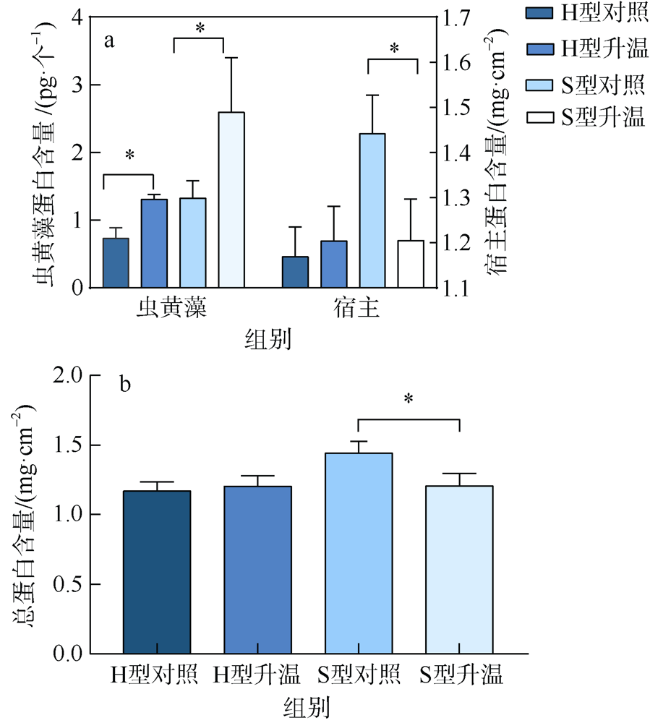
文章研究了温度升高对一种造礁石珊瑚——丛生盔形珊瑚(Galaxea fascicularis)的两种形态型(H和S型)代谢和共生藻光合生理的影响。研究显示H型和S型对升温的生理响应差异明显: 在虫黄藻水平上, H型丢失了大量的虫黄藻, 减轻了共生体呼吸代谢的负荷; S型中虫黄藻数目尚可维持共生体呼吸代谢的需求, 同时提高叶绿素b含量增强光的吸收。在宿主水平上, H型充分利用虫黄藻输送的营养物质, 并通过异养摄食补偿能量消耗; S型宿主所储存的营养物质可以协助共生体适应热环境。结果表明, 丛生盔形珊瑚两种形态型通过不同的能量利用方式适应环境的变化, H型倾向于维持共生体的代谢平衡, 而S型倾向于提高共生藻的能量分配。
何茜 , 俞晓磊 , 梁宇娴 , 张喆 , 黄晖 , 周伟华 , 袁翔城 . 升温对丛生盔形珊瑚两种形态型代谢和共生藻光合生理的影响[J]. 热带海洋学报, 2022 , 41(5) : 133 -140 . DOI: 10.11978/2021078
The effects of warming on the metabolism and photosynthetic physiology of symibiodiniaceae of two morphological types (H and S) of a reef-building coral, Galaxea fascicularis, were studied. The results showed that the physiological responses of H-type and S-type to warming were clearly different. At the symibiodiniaceae level, H-type lost a large number of symibiodiniaceae, and reduced the burden of respiratory metabolism of symbionts. In S-type, the number of zooxanthellae could still maintain the needs of symbionts for respiratory metabolism, while increasing the content of chlorophyll b to enhance light absorption. At the host level, the H-type made full use of nutrients delivered by zooxanthellae and compensated for energy consumption through heterotrophic feeding. The nutrients stored by the S-type host could assist the symbionts to adapt to the thermal environment. Galaxea fascicularis adapted itself to environmental changes through different energy utilization modes, i.e., H-type tended to maintain the metabolic balance of the symbiont, while S-type intended to improve the energy distribution of symbiotic algae.
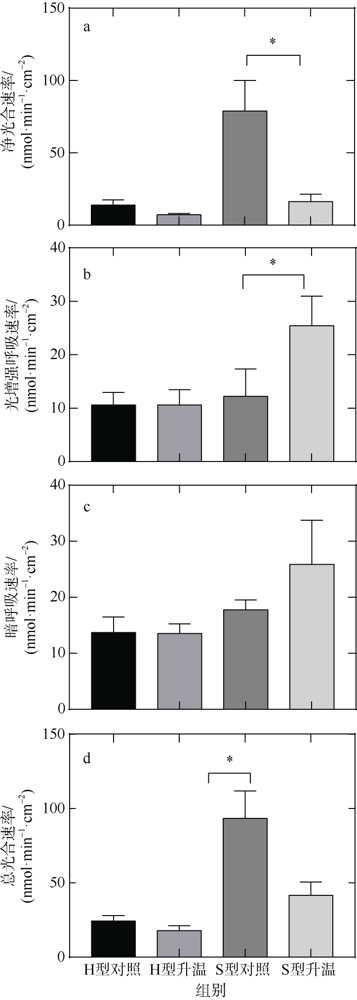
图1 升温对丛生盔形珊瑚H型和S型净光合呼吸速率(a)、光增强呼吸速率(b)、暗呼吸速率(c)和总光合速率(d)的影响*表示差异显著(p<0.05) Fig. 1 The effect of warming on photosynthesis and respiration metabolism of coral. (a) net photosynthetic rate (Pn); (b) light-enhanced respiration rate (RL); (c) dark respiration rate (RD); (d) total photosynthetic rate (Pg). * indicates significant difference (p < 0.05) |
| [1] |
匡志远, 宋振亚, 董昌明, 2020. 基于机器学习订正模型的未来百年全球海表温度预估研究[J]. 气候变化研究快报, 9(4): 270-284.
|
| [2] |
梁宇娴, 俞晓磊, 郭亚娟, 等, 2020. 3种传统方法对不同珊瑚表面积测量的适用性及其校准方法——以3D扫描技术为基准[J]. 热带海洋学报, 39(1): 85-93.
|
| [3] |
吴英, 2018. 丛生盔形珊瑚(Galaxea fascicularis)两种形态型的结构特征与生长特性的比较分析[D]. 海口: 海南大学.
|
| [4] |
俞晓磊, 江雷, 罗勇, 等, 2019. 异养营养对丛生盔形珊瑚代谢及共生藻光合生理的影响[J]. 海洋科学, 43(12): 81-88.
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}