
核酸指标对桡足类安氏伪镖水蚤摄食率、生长率及产卵率的指示作用
作者简介:何学佳(1978—), 女, 贵州省遵义市人, 副研究员, 博士, 主要从事浮游动物生态化学计量学和营养生理生态方面的研究。E-mail: thexuejia@jnu.edu.cn
收稿日期: 2017-05-12
要求修回日期: 2017-06-26
网络出版日期: 2018-04-11
基金资助
国家自然科学基金项目(41276153、41176087)
国家基金委-广东联合重点基金(U1301235)
中国科学院海洋生物资源可持续利用重点实验室开放基金项目(LMB111008)
Nucleic acid proxies for ingestion rate, growth and reproduction in copepod Pseudodiaptomus annandalei
Received date: 2017-05-12
Request revised date: 2017-06-26
Online published: 2018-04-11
Supported by
National Natural Science Foundation of China (41276153, 41176087)
Joint Project of National Nature Science Foundation of China-Guangdong (U1301235)
Open Fund Project of Key Laboratory of Sustainable Utilization of Marine Biological Resources of Chinese Academy of Sciences (LMB111008)
Copyright
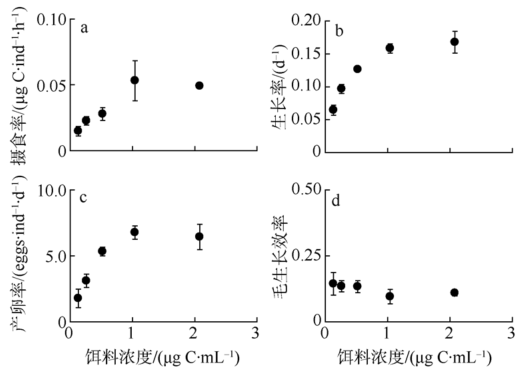
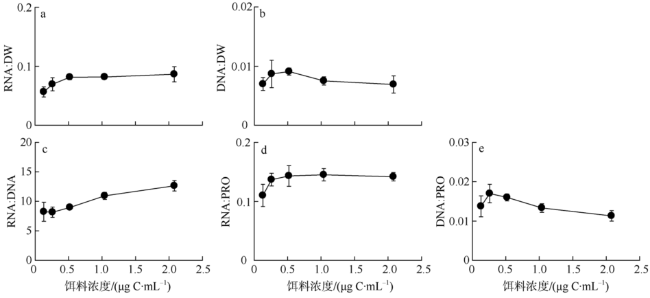
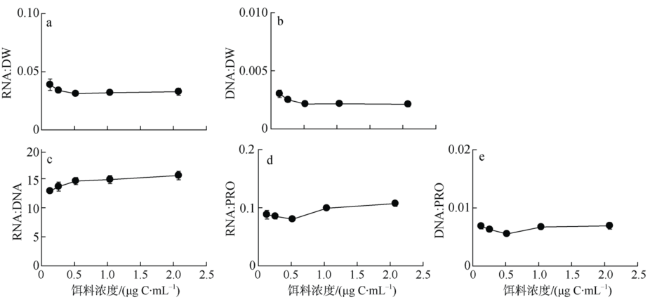
核酸指标可表征桡足类营养及生长状态, 然而多物种和多指标比较研究的相对缺乏却限制了现场海区调查中对核酸标志物的广泛应用。本研究测定了系列饵料浓度(0.2~2.0μg C·mL-1)喂食的安氏伪镖水蚤Pseudodiaptomus annandalei的桡足幼体Ⅱ期个体的摄食率、生长率及成体产卵率, 并就生化组成, 包括RNA含量、DNA含量、蛋白含量(以下简写为PRO)、RNA:DNA、RNA:PRO、DNA:PRO及元素组成(N含量、P含量、C:N 和C:P比值)进行定量分析和计算。摄食率、生长率、产卵率及桡足幼体的RNA相关的核酸指标随饵料浓度的变化趋势均符合双曲线模型, 而雌性成体中仅RNA:DNA比值符合此模型。对RNA相关指标与各生理参数的相关性分析显示, 桡足幼体的RNA相关指标均与摄食率及生长率呈显著线性相关性, 显著程度的顺序为RNA:DNA>RNA:PRO>RNA含量; 雌性成体中RNA:DNA及RNA:PRO比值与产卵率显著相关, 前者的相关性远较后者显著。此结果表明, 在常用的现场浮游桡足类调查的标志比值(RNA:DNA比值或RNA含量)之外, RNA:PRO比值亦可作为指示桡足类生长状态的标志比值, 但是当指示体生长时, 桡足类必须处于同一生长阶段。RNA相关指标与饵料浓度之间稳固的双曲线相关性提示我们, RNA相关指标可帮助评估桡足类的饵料环境。
何学佳 , 杨华明 , 吕柏东 , 尹健强 . 核酸指标对桡足类安氏伪镖水蚤摄食率、生长率及产卵率的指示作用[J]. 热带海洋学报, 2018 , 37(2) : 26 -35 . DOI: 10.11978/2017053
Nucleic acid indices can indicate nutrition status and growth of copepods; yet, lack of comparison among different phylogenetic groups and indices constrains application of nucleic acid proxies in field investigation. In this study, ingestion rate, growth rate of copepodid at Stage Ⅱ and egg production rate were studied in brackish copepod Pseudodiaptomus annandalei at different food levels (0.2~2.0 μg C·mL-1). Biochemical indices including mass specific RNA content, DNA content, RNA:DNA, and RNA: protein ratios, as well as elemental composition (mass specific phosphorus content, nitrogen content, atomic C:P, and C:N ratios ), were also measured and described. Ingestion rate, growth rate, egg production rate, and RNA-related indices in copepodids increased with increasing food level hyperbolically, so did RNA:DNA ratio in female adults. Ingestion and growth rates were significantly correlated with RNA-related indices in copepodids in a linear manner and the strength of correlation ranked as RNA:DNA>RNA:PRO>RNA content. Egg production rate, however, was correlated with RNA:DNA and RNA:PRO ratios in female adults with high and moderate significance, respectively. Our results suggested that RNA:PRO ratio is suitable to be an indicator of growth and reproduction rate in copepod besides RNA:DNA ratio and RNA content, which have been applied in field investigation. However, RNA:DNA as proxy of somatic growth should be confined to the same-aged copepod. In addition, the finding of hyperbolic correlation between RNA-related indices and food concentration implied that they may help prediction of food environment for copepod, but may be confined to food levels at unsaturated phase of functional responses of variables.
Key words: nucleic acid indicators; copepod; ingestion; somatic growth; reproduction
Fig. 1 Physiological variables of P. annandalei in response to food concentration图1 不同饵料浓度条件下安氏伪镖水蚤桡足Ⅱ期个体及雌性成体的生长代谢指标 |
Fig. 2 Biochemical variables of P. annandalei at copedite Stage Ⅱ in reponse to food concentration图2 不同饵料浓度条件下安氏伪镖水蚤桡足Ⅱ期个体的生化组分 |
Fig. 3 Biochemical variables of female P. annandalei in response to food concentration图3 不同饵料浓度条件下安氏伪镖水蚤桡足雌性成体的生化组分 |
Tab. 1 Analysis of functional responses of physiological variables, biochemical and elemental compositions to food concentration in P. annandalei using a hyperbolic equation表1 安氏伪镖水蚤的生理参数、生化及元素组成对饵料浓度的响应的双曲线模型[y=ymax(1-e-b(f+c))]回归分析 |
| 生长阶段 | 指标 | ymax | b | c | r | p |
|---|---|---|---|---|---|---|
| 桡足幼体 | 摄食率 | 0.053±0.006 | 2.04±0.62 | 0 | 0.95 | 0.012 |
| 生长率 | 0.16±0.0053 | 3.3±0.34 | 0 | 0.99 | 0.001 | |
| RNA:干重 | 0.083±0.0019 | 8.1±0.85 | 0 | 0.97 | 0.0054 | |
| RNA:PRO | 0.14±0.0008 | 11±0.37 | 0 | 0.99 | <0.001 | |
| RNA:DNA | 15±3.3 | 0.56±0.38 | 1.1±0.56 | 0.96 | <0.01 | |
| 雌性成体 | 产卵率 | 6.8±0.34 | 2.6±0.38 | 0 | 0.99 | <0.01 |
| RNA:DNA | 15±0.26 | 2.6±0.82 | 0.56±0.20 | 0.97 | <0.001 | |
| C:P | 434±15 | 7.7±1.2 | 0 | 0.95 | 0.015 |
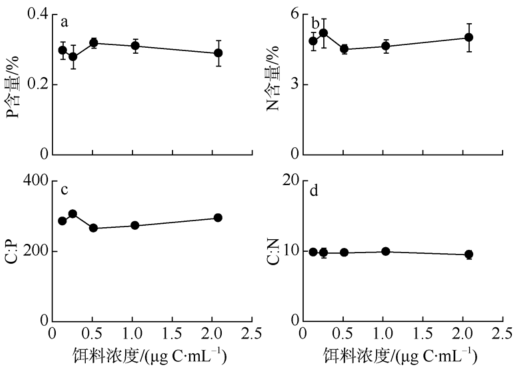
Fig. 4 Elemental composition of P. annandalei at copepodite StageⅡin response to food concentration图4 不同饵料浓度条件下安氏伪镖水蚤桡足Ⅱ期幼体的元素组成 |
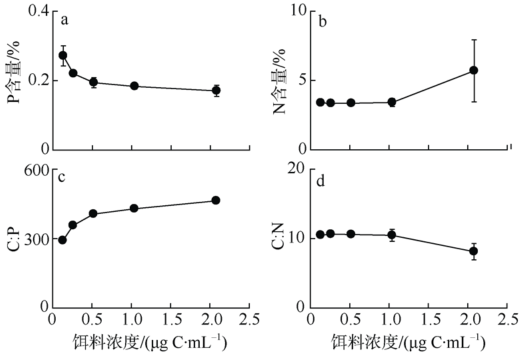
Fig. 5 Elemental composition of female P. annandalei in response to food concentration图5 不同饵料浓度条件下安氏伪镖水蚤雌性成体的元素组成 |
Tab. 2 Analysis of correlations between physiological parameters (ingestion rate, growth rate, and egg production rate) and nucleic acid indices as well as elemental compositions in P. annandalei at different food levels表2 安氏伪镖水蚤蚤体生理指标(摄食率、生长率和产卵率)与核酸指标及元素组成的相关性分析 |
| 生长阶段 | 生理指标 | 核酸指标和元素组成 | Pearson分析 | y=ax+b | ||
|---|---|---|---|---|---|---|
| p | r | a | b | |||
| 桡足Ⅱ期 | 摄食率 | RNA︰干重 | 0.018* | 0.725 | 0.454 | 0.065 |
| RNA︰DNA | 0.000** | 0.953 | 114.8 | 6.956 | ||
| RNA︰PRO | 0.003** | 0.836 | 3.815 | -0.029 | ||
| P含量/% | 0.509 | 0.237 | -0.443 | 0.320 | ||
| N含量/% | 0.797 | 0.093 | 1.05 | 3.957 | ||
| C:P | 0.510 | 0.237 | 155.1 | 103.5 | ||
| C:N | 0.857 | 0.066 | -1.461 | 8.327 | ||
| 桡足Ⅱ期 | 生长率 | RNA︰干重 | 0.005** | 0.688 | 0.165 | 0.074 |
| RNA︰DNA | 0.000** | 0.814 | 39.60 | 4.942 | ||
| RNA︰PRO | 0.004** | 0.702 | 1.118 | -0.064 | ||
| P含量/% | 0.419 | 0.225 | 0.158 | 0.281 | ||
| N含量/% | 0.307 | 0.238 | 1.212 | 3.812 | ||
| C:P | 0.503 | 0.187 | -48.443 | 116.13 | ||
| C:N | 0.528 | 0.178 | -1.515 | 8.514 | ||
| 雌性成体 | 产卵率 | RNA︰干重 | 0.076 | 0.636 | 0.039 | -0.001 |
| RNA︰DNA | 0.000** | 0.851 | 0.511 | 11.71 | ||
| RNA︰PRO | 0.027* | 0.565 | 3.815 | -0.029 | ||
| P含量/% | 0.000** | 0.891 | -0.019 | 0.303 | ||
| N含量/% | 0.193 | 0.356 | 0.244 | 2.593 | ||
| C:P | 0.000** | 0.889 | 12.29 | 88.81 | ||
| C:N | 0.132 | 0.408 | -0.395 | 11.79 | ||
注: 回归分析中, y为核酸指标或化学计量学特征值, x为生理参数; *指显著, **指极显著 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
NAKATA, K, NAKANO, H, KIKUCHI, H, 1994. Relationship between egg productivity and RNA/DNA ratio in Paracalanus sp. in the frontal waters of the Kuroshio. Marine Biology, 119(4): 591-596.
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}