
南海北部陆丰凹陷LF14井中新世古水深变化及沉降特征*
作者简介:薛力园(1989—), 男, 河北省唐山市人, 在读博士研究生, 从事微体古生物学与地层学和古海洋学研究。E-mail: xuealy@cugb.edu.cn
收稿日期: 2017-05-22
要求修回日期: 2017-09-06
网络出版日期: 2018-04-11
基金资助
国家自然科学基金重点项目(91328201)
Miocene evolution of paleo-water depth and subsidence revealed in Well LF14 from Lufeng Sag, northern South China Sea
Received date: 2017-05-22
Request revised date: 2017-09-06
Online published: 2018-04-11
Supported by
National Natural Science Foundation of China (91328201)
Copyright
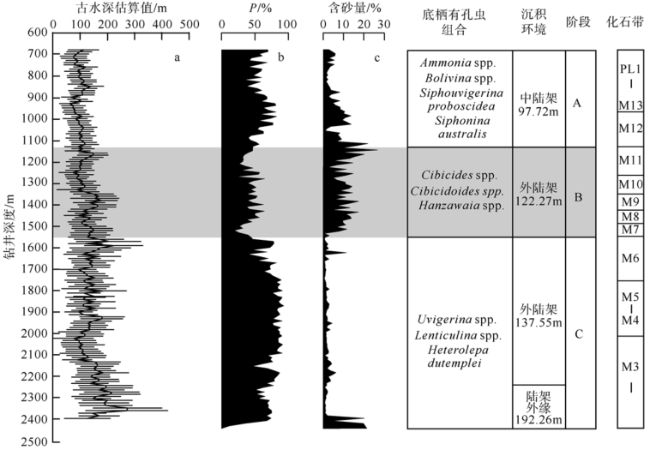
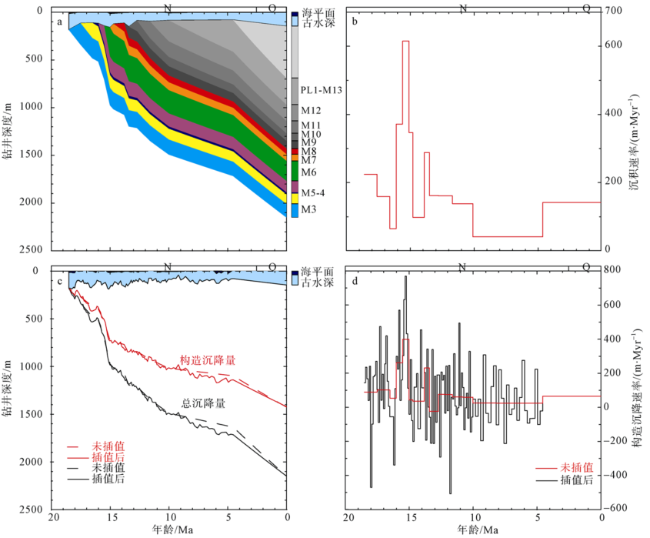
南海北部陆丰凹陷内LF14井记录了早中新世至早上新世的海相沉积地层。古水深重建结果显示研究井位在早中新世突然发生海侵, 水深迅速上升至最大值275m, 随后经数次波动, 整体处于中—外陆架环境, 共记录了5个完整的海侵—海退层序。回剥分析方法重建的LF14井沉降史揭示了研究井位在中中新世(17.5—10Ma)处于快速沉降期, 快速沉降导致的凹陷内沉积物的可容纳空间发育速率高和陆源物质供给充足是造成该阶段沉积速率高的两个重要因素; 晚中新世—早上新世(10—4.53Ma)处于弱沉降期, 推测东沙运动导致凹陷内的沉积物可容纳空间发育速率变小和陆源物质供给减少, 造成该时期内研究井位沉积速率低。最后, 依据定量重建的古水深变化在研究层段识别出一系列短暂存在的构造上升事件。
薛力园 , 丁旋 , 裴人傑 , 万晓樵 . 南海北部陆丰凹陷LF14井中新世古水深变化及沉降特征*[J]. 热带海洋学报, 2018 , 37(2) : 72 -83 . DOI: 10.11978/2017060
Well LF14, drilled in the Lufeng Sag of the northern South China Sea, discloses marine sediment archives ranging from the mid-to-late period of the early Miocene to the early Pliocene. An abrupt rise in paleo-water depth up to 275 m during the early Miocene is recorded at the lowermost part of the well, followed by several fluctuations thereafter. The early Miocene to early Pliocene deposits are interpreted as formed under middle-to-outer shelf environment, and five complete transgressive-regressive sea-level cycles are identified. The results of backstripping calculations indicate a rapid subsidence in the study area during 17.5-10 Ma. The high sedimentation rate in this period may be caused by rapid tectonic subsidence and high terrigenous input. The subsidence rate was low from 10 to 4.53 Ma, characterized by a low sedimentation rate mainly due to less supply of terrigenous materials and less accommodation space caused by the Dongsha Movement. Finally, several short-lived uplift events were identified in the well based on quantitative reconstruction of paleo-water depth.
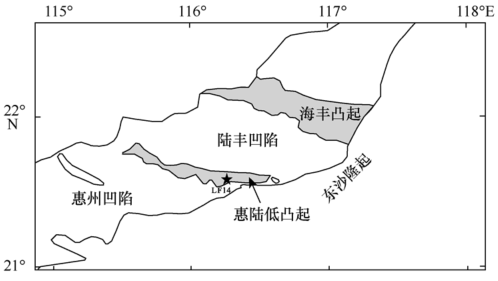
Fig. 1 Location of Well LF14. Modified after Zhang et al (2012)图1 LF14井位置图(据张向涛 等, 2012) |
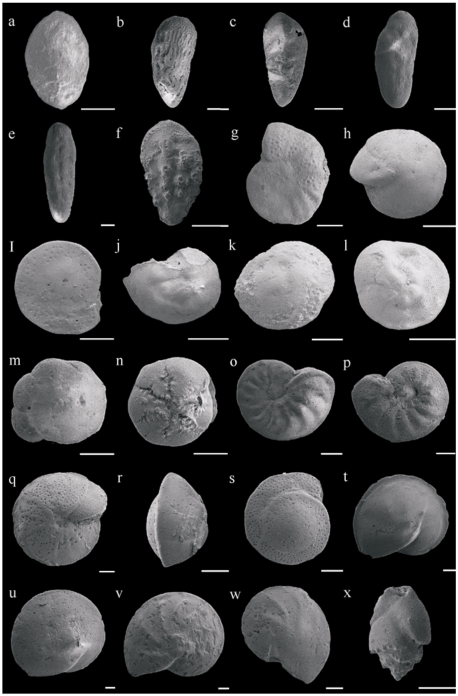
Fig. 2 Some major benthic foraminifera genera/species in Well LF14 (all scale bars are 100μm)图2 LF14井主要底栖有孔虫属种 |
Fig. 3 Some major benthic foraminifera genera/species in Well LF14 (All scale bars are 100μm)图3 LF14井主要底栖有孔虫属种 |
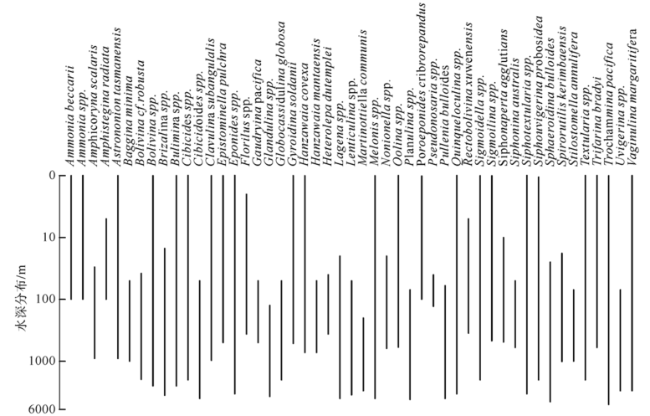
Fig. 4 Water depth range of benthic foraminifera predominant in Well LF14. Bathymetric ranges are based on Van Morle (1988), Wang et al (1988), González-Regalado (1989), Murray (1991, 2006), Zheng et al (2001), Gupta (2003), Pérez-Asensio et al (2012), and “Ocean Biogeographic Information System” (http://www.iobis.org)图4 LF14井底栖有孔虫水深分布范围 |
Tab. 1 Biostratigraphic controls of Well LF14 and global sea level change表1 LF14井生物地层年龄控制点和全球海平面变化 |
| 深度/m | 化石带 | 浮游有孔虫生物事件 (LAD) | 年龄/Ma | 全球海平面变化/m |
|---|---|---|---|---|
| 685±5 | PL1 | Sphaeroidinellopsis kochi | 4.53 | 9.6 |
| 975±5 | M13/M12 | Globoquadrina dehiscens | 10 | -5.19 |
| 1135±5 | M12/M11 | Globorotalia siakensis | 11.6 | 17.79 |
| 1265±5 | M11/M10 | Globigerinoides subquadratus | 12.7 | 5.48 |
| 1345±5 | M10/M9 | Globorotalia foshi | 13.4 | -1 |
| 1425±5 | M9/M8 | Globorotalia foshi peripheroacuta | 13.8 | -2.13 |
| 1485±5 | M8/M7 | Globorotalia foshi peripheroronda | 14.7 | -9.4 |
| 1555±5 | M7/M6 | Globigerinoides sicanus | 15 | -12 |
| 1755±5 | M6/M5 | Globigerinatella insueta | 15.5 | 15 |
| 1885±5 | M5-4 | Globorotalia birnageae | 16.00 | 0.93 |
| 1905±5 | M5-4 | Globigerinoides parawoodi | 16.50 | 3.1 |
| 2005±5 | M4/M3 | Catapsydrax dissimilis | 17.5 | 4.15 |
注: LAD(last appearance datum)指末现面。年龄值参考自秦国权(1996), 全球海平面变化参考Miller等(2005) |
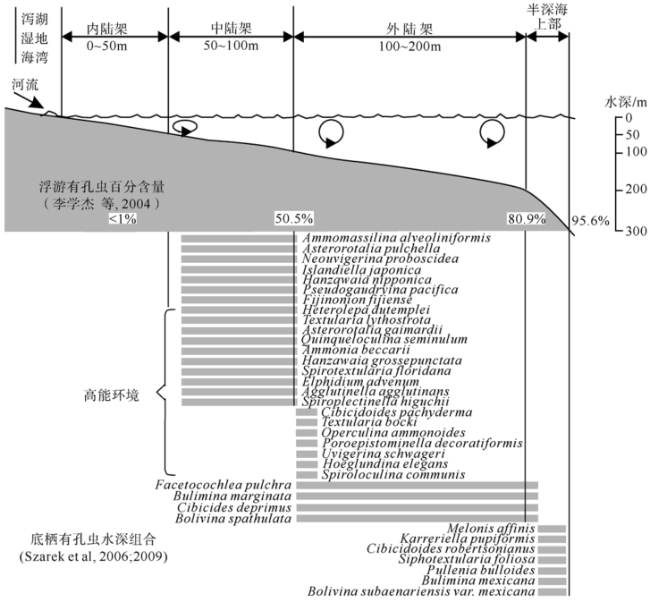
Fig. 5 Summary of benthic foraminiferal assemblages observed across the continental slope of the Sunda Shelf in the southern South China Sea图5 南海南部Sunda陆架陆坡区底栖有孔虫组合分布 |
Fig. 6 Estimated paleodepth curves for Well LF14图6 LF14井古水深估算曲线 |
Fig. 7 Burial history (a), sedimentation rate (b), subsidence chart (c), and tectonic subsidence rate (d) in Well LF14 (N, Neocene; Q, Quaternary)图7 LF14井埋藏史(a)、沉积速率(b)、沉降史(c)和构造沉降速率(d)变化 |
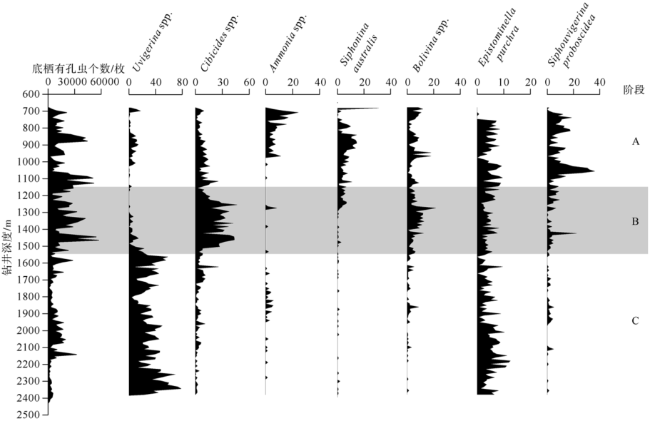
Fig. 8 Number of benthic foraminifera and relative abundance of water depth markers in Well LF14图8 LF14井底栖有孔虫个数(枚)和水深指示种百分含量(%)变化曲线 |
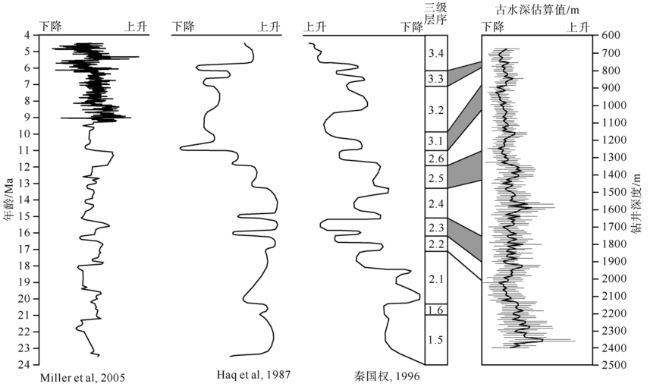
Fig. 9 Comparison between the regional and global sea level curves and the paleo depth of studied interval from Well LF14. Modified according to Haq et al (1987), Miller et al (2005) and Wang et al (2009)图9 LF14井古水深与区域和全球海平面变化曲线对比 |
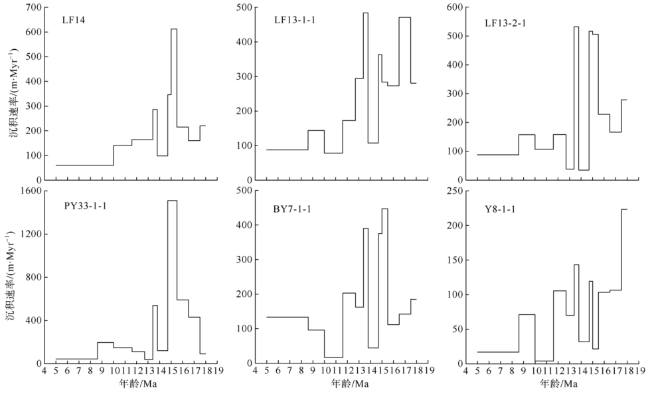
Fig. 10 Comparison of Miocene sedimentation rate between Well LF14 and other wells from the northern South China Sea. Data for Well LF13-1-1, LF13-2-1, PY33-1-1, BY7-1-1, and Y8-1-1 are from Dong (1996)图10 LF14井与南海北部其他井位中新世沉积速率对比 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
湖北省地质科学研究所, 河南省地质局, 湖北省地质局, 等, 1978. 中南地区古生物图册(四): 微体化石部分[M]. 北京: 地质出版社.
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
WEI KUISHENG, CUI HANYUN, YE SHUFEN, et al, 2001. High-precision sequence stratigraphy in Qiongdongnan Basin[J]. Earth Science-Journal of China University of Geosciences, 26(1): 59-66 (in Chinese with English abstract).
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
VAN DER ZWAAN G J,
|
| [65] |
VAN HINSBERGEN D J J,
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}