
斑节对虾3个野生群体遗传多样性的微卫星标记分析
作者简介:赵志英(1968—), 男, 海南东方市人, 博士, 副研究员, 从事水生动物遗传育种及病害防治研究。E-mail: zhaozhying@163.com
收稿日期: 2017-07-30
要求修回日期: 2017-10-17
网络出版日期: 2018-05-03
基金资助
海南省应用技术研发与示范推广专项(ZDXM2015025)
Analysis of genetic diversity among three wild populations of Penaeus monodon using microsatellite marker
Received date: 2017-07-30
Request revised date: 2017-10-17
Online published: 2018-05-03
Supported by
Application Technology R & D and Demonstration Project of Hainan Province (ZDXM2015025)
Copyright
为探究不同地理条件下斑节对虾天然群体的遗传多样性和遗传分化水平, 选用14对简单重复序列(simple sequence repeat, SSR)特异引物对广东湛江、海南三亚、海南琼海的斑节对虾天然群体共计90尾个体进行微卫星标记分析, 每对引物检测出的等位基因数为2~4不等, 平均2.7。3个群体(湛江、三亚、琼海)平均有效等位基因分别为1.9081、1.9715和2.0185, 平均多态信息含量分别为0.3864、0.3926和0.4078, 总的平均期望杂合度(0.4605 )明显高于观测杂合度(0.4046), 表明3个群体遗传多样性水平为中等, 且存在杂合度缺失的现象。种群分化指数和遗传距离分析表明, 在不同群体间已产生了遗传分化, 但是分化较小; 三亚群体和琼海群体之间的亲缘关系最近, 而与湛江群体的亲缘关系最远。分子方差分析表明, 群体间的遗传变异指数为8%, 群体内个体间的遗传变异指数为92%, 基因分化主要发生在群体内, 而不是群体之间。
赵志英 , 梁丽运 , 白丽蓉 . 斑节对虾3个野生群体遗传多样性的微卫星标记分析[J]. 热带海洋学报, 2018 , 37(3) : 65 -72 . DOI: 10.11978/2017084
Genetic diversity and genetic differentiation among 90 individuals of Penaeus monodon from three wild populations (Zhanjiang, Sanya, and Qionghai) were investigated by using 14 pairs of simple sequence repeat (SSR) specific primers. The number of alleles detected by each pair of primers ranged from 2 to 4, with the average of 2.7. The mean polymorphism information contents of the three populations were 0.3864, 0.3926, and 0.4078, respectively; and the mean effective numbers of alleles were 1.9081, 1.9715 and 2.0185, respectively. The average expected heterozygosity (0.4605) of the three populations was significantly higher than the observed heterozygosity (0.40469), indicating moderate polymorphism and the existence of heterozygosity deficiency among the three populations. The results of genetic distances and genetic similarity showed that the genetic differentiation among the three populations was low; genetic variation occurred mainly within populations, not among populations. The genetic relationship of Sanya and Qionghai populations was the closest, while that of Sanya and Zhanjiang populations was the farthest. Analysis of molecular variance (AMOVA) revealed that most of the genetic variation resided within populations (92%), and less, among populations (8%), suggesting rich genetic diversity level within populations.
Tab. 1 Fourteen pairs of effective microsatellites primers sequence and their annealing temperature表1 14对有效扩增的微卫星引物及退火温度 |
| 微卫星位点 | GenBank序列号 | 引物序列(5′-3′) | 退火温度/℃ |
|---|---|---|---|
| DTLPM218 | AY 189727 | F: ATTCCGCAATATATCGGTTTCC | 57 |
| R: AATGTTTCCATTTCATGCTTCG | |||
| DTLPM303 | AY 188984 | F: TGCCTTGTATTTTGACGATCAG | 56 |
| R: TTGGAGTAGCAACAGCGGTA | |||
| DTLPM402 | AY 188987 | F: CCACTCTAACTCCGCCAGTC | 54 |
| R: TCCCTACCCCACTATCATCG | |||
| DTLPM 120 | AY188978 | F: TTATCCGTATAGCCGCGTTATC | 52 |
| R: TTACAGGACCTGCATTTGTGTC | |||
| DTLPM 104 | AY188974 | F: AGGACCTGCATTTGTGTCG | 52 |
| R: ATGGCGAGACAAGGTTCG | |||
| BC52 | AF097631 | F: AAGCAGATAGACCGTCAG | 52 |
| R: AATCCTGCAGAGCAATAC | |||
| TUZXPM49 | AF077568 | F: ATCTGACAGGGCACCATAC | 54 |
| R: AGTCGAGTCTTGAATAAGCG | |||
| PM138 | P1-AY500853 | F: ACGGAGTGGGTAGAGACATA | 56 |
| R: ACAAGCGAAGTGAAGAGG | |||
| PM205 | AY500854 | F: AGGAATGATGGGAGGGAAAG | 56 |
| R: AAGCTCAGGCAAGCGTGTAT | |||
| PM528 | AY500855 | F: GTGTTATTTTCCACGGGTGC | 56 |
| R: GCTGCAGGAAGTGTAGGGAG | |||
| PM1713 | AY500858 | F: GTTGCGACGGGTTGATTC | 54 |
| R: TTTATGGCTATGGCTGACAC | |||
| PM2345 | AY500860 | F: GATATTTCAAGGAATGCTCG | 54 |
| R: TAATTCGTGCCTTACCTCAT | |||
| PM3852 | AY500862 | F: TAATGGGCGTAAGTCTTCGG | 56 |
| R: TGAAAGGAGTCGGGATATGC | |||
| PM4793 | AY500867 | F: CTTCTAGCGCCATTTCAAGG | 56 |
| R: TCCTTCCAGTGTTCGGAGTT |
Tab. 2 Allelic frequency of 14 microsatellite loci in three populations of P. monodon表2 14个微卫星位点在3个斑节对虾群体中的等位基因频率 |
| 位点 | 等位基因 | 群体 | 位点 | 等位基因 | 群体 | ||||
|---|---|---|---|---|---|---|---|---|---|
| 琼海 | 三亚 | 湛江 | 琼海 | 三亚 | 湛江 | ||||
| PM1384 | A | 0.500 | 0.4667 | 0.4167 | PM1383 | A | 0.7333 | 0.2667 | 0.4667 |
| B | 0.0667 | 0.0167 | 0.0333 | B | 0.1500 | 0.4000 | 0.2333 | ||
| C | 0.400 | 0.3000 | 0.4833 | C | 0.1167 | 0.3333 | 0.3000 | ||
| D | 0.0333 | 0.2167 | 0.0667 | ||||||
| TUZXPM49 | A | 0.0500 | 0.0333 | 0.0167 | PM1382 | A | 0.3000 | 0.3000 | 0.4667 |
| B | 0.5167 | 0.5667 | 0.4667 | B | 0.3000 | 0.0833 | 0.0500 | ||
| C | 0.4333 | 0.4000 | 0.5167 | C | 0.4000 | 0.6167 | 0.4833 | ||
| PM1713 | A | 0.7333 | 0.6500 | 0.7333 | DTLPM104 | A | 0.2333 | 0.1000 | 0.0500 |
| B | 0.2667 | 0.3500 | 0.2667 | B | 0.6833 | 0.7833 | 0.6333 | ||
| C | 0.0833 | 0.1167 | 0.3167 | ||||||
| BC52 | A | 0.3167 | 0.5833 | 0.0667 | PM1381 | A | 0.6500 | 0.4667 | 0.3833 |
| B | 0.6833 | .4167 | 0.9333 | B | 0.3500 | 0.5333 | 0.6167 | ||
| PM528 | A | 0.3167 | 0.1500 | 0.3167 | PM138 | A | 0.0500 | — | 0.0667 |
| B | 0.5333 | 0.6333 | 0.5167 | B | 0.6833 | 0.9333 | 0.8333 | ||
| C | 0.1500 | 0.2167 | 0.1667 | C | 0.2667 | 0.0667 | 0.1000 | ||
| PM2345 | A | 0.0833 | 0.0667 | 0.0167 | PM3852 | A | 0.1667 | 0.1833 | 0.1333 |
| B | 0.8667 | 0.9333 | 0.9333 | B | 0.5833 | 0.6500 | 0.6333 | ||
| C | 0.0500 | — | 0.0500 | C | 0.2500 | 0.1667 | 0.2333 | ||
| PM205 | A | 0.6167 | 0.6167 | 0.6667 | PM4793 | A | 0.5167 | 0.4333 | 0.7500 |
| B | 0.3833 | 0.3833 | 0.3333 | B | 0.4833 | 0.5667 | 0.2500 | ||
注: 每个位点的等位基因均从A开始。 |
Tab. 3 Genetic variation of three populations (Qionghai, Sanya, and Zhanjiang) of P. monodon表3 斑节对虾3个地理群体(琼海/三亚/湛江)的遗传变异 |
| 位点 | 等位 基因数 | 有效等位 基因数 | 观察杂合度 | 期望杂合度 | HW平衡偏离指数 | HW 平衡检测 | 多态信息含量 |
|---|---|---|---|---|---|---|---|
| DTLPM120 | 4/4/4 | 2.4064/2.8169/2.4226 | 1.000/0.9333/0.9333 | 0.5844/0.6450/0.5872 | 0.7111/0.4470/0.5894 | **/**/* | 0.5804/0.6352/0.5842 |
| TUZXPM49 | 3/3/3 | 2.1871/2.0737/2.0619 | 0.4667/0.6667/0.4000 | 0.5428/0.5178/0.5150 | -0.1402/0.2876/-0.2233 | ns/ns/ns | 0.4212/0.4162/0.3985 |
| PM1713 | 2/2/2 | 1.6423/1.8349/1.6423 | 0.2667/0.5667/0.4667 | 0.3911/0.4550/0.3911 | -0.3181/0.2455/0.1933 | ns/ns/ns | 0.3146/0.3515/0.3146 |
| BC52 | 2/2/2 | 1.7630/1.9459/1.1421 | 0.5667/0.3667/0.1333 | 0.4328/0.4861/0.1244 | 0.3094/-0.2456/-0.0715 | ns/ns/ns | 0.3391/0.3668/0.1168 |
| DTLPM402 | 3/3/3 | 1.7425/2.9221/2.7607 | 0.3333/0.3000/0.4000 | 0.4261/0.6578/0.6378 | -0.2178/-0.5439/-0.3728 | ns/**/* | 0.4013/0.5995/0.6043 |
| DTLPM303 | 3/3/3 | 2.9412/2.0955/2.2032 | 0.5000/0.6333/0.4000 | 0.6600/0.5228/0.5461 | -0.2424/0.2114/-0.2675 | ns/ns/ns | 0.51/0.5162/0.5439 |
| DTLPM104 | 3/3/3 | 1.8927/1.5693/1.9846 | 0.4667/0.4333/0.4667 | 0.4717/0.3628/0.4961 | -0.011/0.1943/-0.0593 | ns/ns/ns | 0.4146/0.3338/0.4137 |
| DTLPM218 | 2/2/2 | 1.8349/1.9912/1.8967 | 0.4333/0.4667/0.2333 | 0.4550/0.4978/0.4728 | -0.0478/-0.0625/-0.5066 | ns/ns/** | 0.3515/0.3739/0.3857 |
| PM528 | 3/3/3 | 2.4557/2.1251/2.5316 | 0.4000/0.3000/0.4333 | 0.5928/0.5294/0.6050 | -0.3252/-0.4333/-0.2838 | */**/ns | 0.5231/0.4738/0.5365 |
| PM205 | 2/2/2 | 1.8967/1.8967/1.8000 | 0.3667/0.6333/0.6000 | 0.4728/0.4728/0.4444 | -0.2244/0.3395/0.3501 | ns/ns/ns | 0.3610/0.3610/0.3457 |
| PM138 | 3/2/3 | 1.8499/1.1421/1.4107 | 0.4333/0.0000/0.1333 | 0.4594/0.1244/0.2911 | -0.0568/-1.000/-0.5421 | ns/**/** | 0.3902/0.1168/0.2711 |
| PM3852 | 3/3/3 | 2.3226/2.0666/2.1127 | 0.3667/0.2333/0.2333 | 0.5694/0.5161/0.5267 | -0.3559/-0.5480/-0.5571 | */**/** | 0.5025/0.4642/0.4688 |
| PM2345 | 3/2/3 | 1.3148/1.1421/1.1443 | 0.1667/0.0667/0.0667 | 0.2394/0.1244/0.1261 | -0.3037/-0.4638/-0.4711 | **/*/** | 0.2252/0.1168/0.1213 |
| PM4793 | 2/2/2 | 1.9978/1.9651/1.6000 | 0.1000/0.5333/0.2333 | 0.4994/0.4911/0.3750 | -0.7998/0.0860/-0.3787 | **/ns/* | 0.3747/0.3705/0.3047 |
| 平均 | 2.7143/ 2.5714/ 2.7143 | 2.0185/1.9715/1.9081 | 0.4214/0.4357/0.3667 | 0.4857/0.4575/0.4385 | -0.2926/-0.2809/-0.2817 | ns/ns/ns | 0.4078/0.3926/0.3864 |
注: ns表示差异不显著(P>0.05); *表示差异显著(P<0.05); **表示差异极显著(P<0.01)。 |
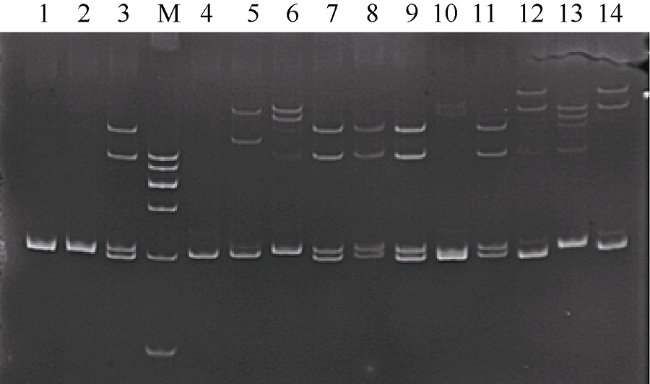
Fig. 1 Electrophoretic results of microsatellite locus PM205 in different individuals of P. monodon图1 微卫星PM205检测部分斑节对虾个体的电泳结果 |
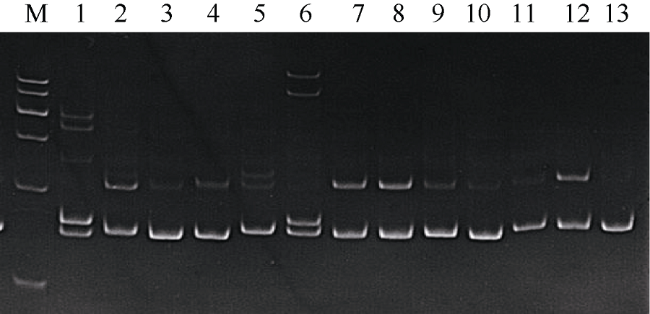
Fig. 2 Electrophoretic results of microsatellite locus PM1713 in different individuals P. monodon图2 微卫星PM1713检测部分斑节对虾个体的电泳结果 |
Tab. 4 Genetic distance and genetic similar index in the three populations of P. monodon表4 斑节对虾3个群体的遗传距离及相似性指数 |
| 琼海 | 三亚 | 湛江 | |
|---|---|---|---|
| 琼海 | 0.9569 | 0.9461 | |
| 三亚 | 0.0440 | 0.9355 | |
| 湛江 | 0.0554 | 0.0667 |
注: 对角线以上数据为相似性指数, 对角线以下数据为遗传距离。 |
Tab. 5 F-value and gene flow of 14 microsatellite loci in the three populations of P. monodon表5 14 个微卫星位点在3个群体斑节对虾中的F值和基因流(Nm) |
| 微卫星位点 | 近交系数 | 遗传分化系数 | 基因流 | 微卫星位点 | 近交系数 | 遗传分化系数 | 基因流 |
|---|---|---|---|---|---|---|---|
| DTLPM120 | -0.5780 | 0.0219 | 11.1477 | DTLPM402 | 0.4197 | 0.0887 | 2.5679 |
| TUZXPM49 | 0.0070 | 0.0079 | 31.3235 | DTLPM218 | 0.2050 | 0.0496 | 4.7873 |
| PM1713 | -0.0507 | 0.0074 | 33.4050 | DTLPM303 | 0.1131 | 0.0438 | 5.4533 |
| BC52 | -0.0224 | 0.2038 | 0.9768 | DTLPM104 | -0.0271 | 0.0442 | 5.4104 |
| PM528 | 0.3438 | 0.0165 | 14.9471 | PM3852 | 0.4831 | 0.0047 | 53.0854 |
| PM205 | -0.1511 | 0.0024 | 104.2500 | PM2345 | 0.3878 | 0.0142 | 17.4079 |
| PM138 | 0.3524 | 0.0612 | 3.8352 | PM4793 | 0.3653 | 0.0732 | 3.1675 |
| 平均 | 0.1143 | 0.0456 | 5.2321 |
Tab. 6 Genetic differentiation coefficient in the three populations of P. monodon表6 斑节对虾3个群体的遗传分化指数 |
| 琼海 | 三亚 | 湛江 | |
|---|---|---|---|
| 琼海 | |||
| 三亚 | 0.0332 | ||
| 湛江 | 0.0288 | 0.0419 |
Tab. 7 Analysis on molecular variance of genetic differentiation in the three populations of P. monodon表7 斑节对虾3个群体遗传分化的分子方差分析 |
| 变异来源 | 自由度 | 平方和 | 方差组成 | 变异百分比/% |
|---|---|---|---|---|
| 群体间 | 2 | 55.389 | 0.677 | 8 |
| 群体内 | 87 | 641.800 | 7.377 | 92 |
| 合计 | 89 | 697.189 | 8.054 | 100 |
Fig.3 Dendrogram of three P. monodon populations based on UPGMA clustering analysis图3 UPGMA聚类分析法构建3个群体斑节对虾的谱系关系图 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}