
南海中西部渔场主要渔业生物碳氮稳定同位素特征
作者简介:黄佳兴(1994—), 男, 江西省新余市人, 硕士研究生, 研究方向为渔业资源调查与评估。E-mail: 820756571@qq.com
收稿日期: 2018-04-16
要求修回日期: 2018-07-08
网络出版日期: 2019-01-16
基金资助
国家重点基础研究发展计划(“973”)项目(2014CB441505)
农业农村部财政专项项目(NFZX2013)
中国水产科学研究院基本科研业务费资助(2017HY-ZD0804)
Characteristics of stable carbon and nitrogen isotopes of major fishery organisms in the fishing ground of central western South China Sea
Received date: 2018-04-16
Request revised date: 2018-07-08
Online published: 2019-01-16
Supported by
National Basic Research Program of China (“973”) (2014CB441505)
Financial Fund of the Ministry of Agriculture and Rural Affairs (NFZX2013)
Special Scientific Research Funds for Central Non-profit Institutes, Chinese Academy of Fishery Sciences (2017HY-ZD0804)
Copyright
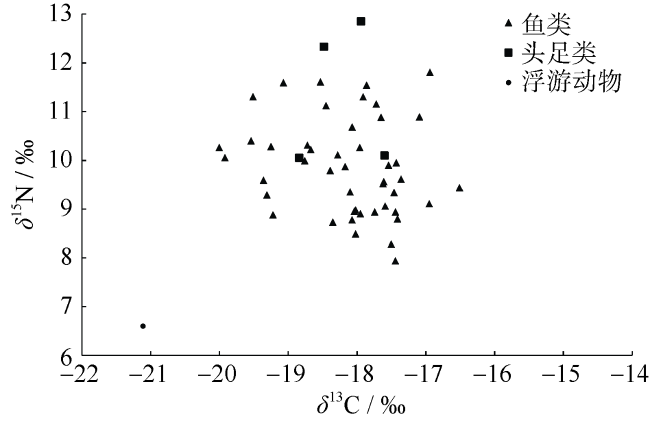
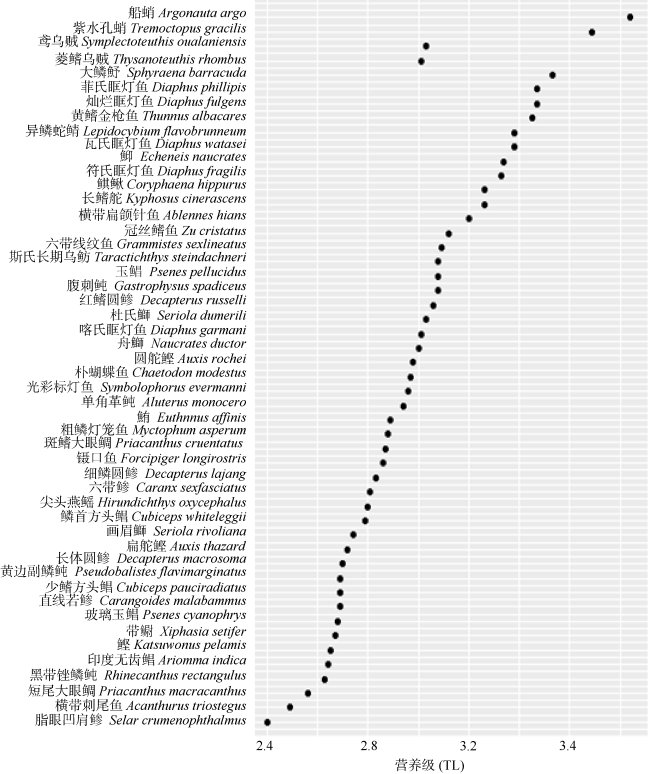
南海中西部渔场是我国南海渔业开发的重点渔场之一。为了解该海域主要渔业生物的营养关系, 应用碳、氮稳定同位素技术测定了该海域主要渔业生物样品的δ13C和δ15N值, 由此构建该海域主要渔业生物的连续营养谱。结果显示, 南海中西部渔场主要渔业生物同位素比值变化幅度较大, 其中鱼类的δ13C和δ15N值范围分别为-20.00‰~-16.51‰和7.94‰~11.81‰; 头足类的δ13C和δ15N值范围分别为-18.84‰~-17.60‰和10.10‰~12.85‰。以浮游动物为基线生物计算各物种相应的营养级, 鱼类处于2.41~3.53, 头足类处于3.03~3.84, 头足类的平均营养级要高于鱼类。通过对不同体长( 胴长)的鸢乌贼Symplectoteuthis oualaniensis、菱鳍乌贼Thysanoteuthis rhombus、红鳍圆鲹Decapterus russelli、细鳞圆鲹Decapterus lajang和黄鳍金枪鱼Thunnus albacares的营养级进行比较分析, 结果发现, 随着体长(胴长)增大其营养级有相应增大的趋势。研究初步建立了南海中西部渔场主要渔业生物营养级的连续营养谱, 旨在为了解该海域食物网结构提供基础资料。
黄佳兴 , 龚玉艳 , 徐姗楠 , 王欢欢 , 张魁 , 张俊 , 陈作志 . 南海中西部渔场主要渔业生物碳氮稳定同位素特征[J]. 热带海洋学报, 2019 , 38(1) : 76 -84 . DOI: 10.11978/2018041
The central western South China Sea is one of the main fishing grounds for fishery development. To understand the trophic relationships of major fishery organisms of the central western South China Sea, stable isotope techniques were used to analyze and determine carbon and nitrogen stable isotope ratios of major fishery biological samples in the area. The trophic levels of the main fishery biota were calculated to construct a continuous trophic spectrum of the main fishery organisms in the area. These results show that the major fishery isotope ratios have a wide range of changes in the central western South China Sea. The δ13C and δ15N values of the fish ranged from -20.00‰ to -16.51‰ and from 7.94‰ to 11.81‰, respectively. The δ13C and δ15N values of the cephalopods ranged from -18.84‰ to -17.60‰ and from 10.10‰ to 12.85‰, respectively. The corresponding trophic levels of each species were calculated using zooplankton as the baseline organism. The trophic level of fish ranged from 2.41 to 3.53, and that of cephalopods ranged from 3.03 to 3.84. Among the organisms, the average trophic level of cephalopods is higher than that of fish. Comparison of trophic levels in different lengths (mantle length) of Symplectoteuthis oualaniensis, Thysanoteuthis rhombus, Decapterus russelli, Decapterus lajang, and Thunnus albacares reveals that the trophic level has a correspondingly increasing trend as body length increases. In this study, we preliminarily established the continuous trophic levels’ spectrum of major fishery bio-nutritional levels in the central and western waters of the central western South China Sea, providing a theoretical basis for the food web structure and fishery resource utilization in the area.
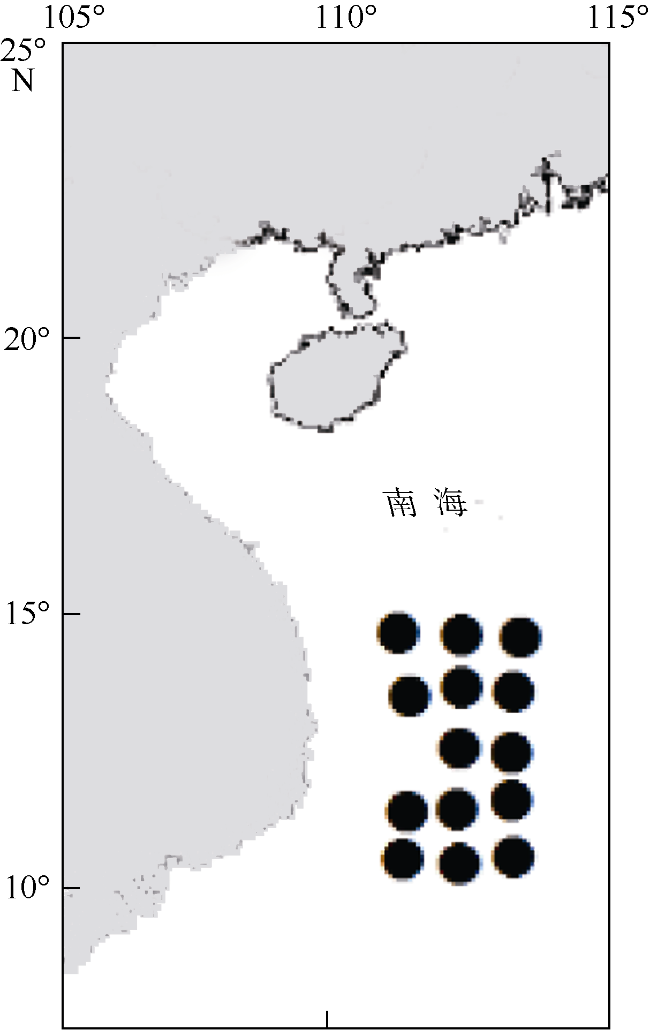
Fig.1 The central western South China Sea, and sampling stations图1 南海中西部采样站点图 |
Tab. 1 The length range, δ13C value, δ15N value, and trophic level (TL) of fishery organisms in the central western South China Sea表1 南海中西部海域渔业生物的体长范围、δ13C值、δ15N值和营养级(TL) |
| 种 类 | 体长范围/mm | δ13C | δ15N | TL | 尾数 | |
|---|---|---|---|---|---|---|
| 鱼类 | 脂眼凹肩鲹Selar crumenophthalmus | 32.1~113.6 | -17.44 | 7.94 | 2.40 | 3 |
| 横带刺尾鱼Acanthurus triostegus | 48.3~52.6 | -17.50 | 8.28 | 2.49 | 2 | |
| 短尾大眼鲷Priacanthus macracanthus | 72.4~78.3 | -18.02 | 8.49 | 2.56 | 2 | |
| 黑带锉鳞鲀Rhinecanthus rectangulus | 28.3 | -18.35 | 8.73 | 2.63 | 1 | |
| 印度无齿鲳Ariomma indica | 147.8 | -18.07 | 8.78 | 2.64 | 1 | |
| 鲣Katsuwonus pelamis | 243.3 | -17.41 | 8.80 | 2.65 | 1 | |
| 带鳚Xiphasia setifer | 378.6 | -19.22 | 8.88 | 2.67 | 1 | |
| 玻璃玉鲳Psenes cyanophrys | 144.6~158.7 | -17.95 | 8.90 | 2.68 | 2 | |
| 直线若鲹Carangoides malabammus | 162.3 | -17.44 | 8.94 | 2.69 | 1 | |
| 少鳍方头鲳Cubiceps pauciradiatus | 99~122 | -17.74 | 8.94 | 2.69 | 7 | |
| 黄边副鳞鲀Pseudobalistes flavimarginatus | 58.9~66.7 | -18.04 | 8.95 | 2.69 | 2 | |
| 长体圆鲹Decapterus macrosoma | 103.3~135.3 | -18.02 | 8.98 | 2.70 | 7 | |
| 扁舵鲣Auxis thazard | 184.6~236.7 | -17.59 | 9.06 | 2.72 | 7 | |
| 细鳞圆鲹Decapterus lajang | 257~300 | -16.51 | 9.44 | 2.83 | 6 | |
| 画眉鰤Seriola rivoliana | 232 | -16.95 | 9.11 | 2.74 | 1 | |
| 鳞首方头鲳Cubiceps whiteleggii | 133.3 | -19.31 | 9.29 | 2.79 | 1 | |
| 尖头燕鳐Hirundichthys oxycephalus | 132.7~203.9 | -17.46 | 9.34 | 2.80 | 5 | |
| 六带鲹Caranx sexfasciatus | 118.7 | -18.10 | 9.35 | 2.81 | 1 | |
| 镊口鱼Forcipiger longirostris | 41.3~54.7 | -17.62 | 9.52 | 2.86 | 2 | |
| 斑鳍大眼鲷Priacanthus cruentatus | 84.3 | -17.61 | 9.56 | 2.87 | 1 | |
| 粗鳞灯笼鱼Myctophum asperum | 57.2~68.7 | -19.36 | 9.59 | 2.88 | 5 | |
| 鮪Euthnnus affinis | 174.3~187.2 | -17.36 | 9.61 | 2.89 | 6 | |
| 单角革鲀Aluterus monocero | 337.6~368.4 | -18.39 | 9.79 | 2.94 | 4 | |
| 光彩标灯鱼Symbolophorus evermanni | 63.2~72.3 | -18.17 | 9.87 | 2.96 | 4 | |
| 红鳍圆鲹Decapterus russelli | 127.3~134.5 | -18.67 | 10.22 | 3.06 | 3 | |
| 朴蝴蝶鱼Chaetodon modestus | 58.4~59.6 | -17.54 | 9.90 | 2.97 | 2 | |
| 圆舵鲣Auxis rochei | 185.3~254.3 | -17.43 | 9.95 | 2.98 | 12 | |
| 舟鰤Naucrates ductor | 168.3~208.4 | -18.76 | 9.99 | 3.00 | 6 | |
| 喀氏眶灯鱼Diaphus garmani | 37.2 | -19.92 | 10.05 | 3.01 | 1 | |
| 杜氏鰤Seriola dumerili | 142.3~147.6 | -18.28 | 10.11 | 3.03 | 2 | |
| 腹刺鲀Gastrophysus spadiceus | 324.4 | -17.96 | 10.26 | 3.08 | 1 | |
| 玉鲳Psenes pellucidus | 44.3~7.2 | -20.00 | 10.26 | 3.08 | 2 | |
| 斯氏长期乌鲂Taractichthys steindachneri | 133.2 | -19.25 | 10.28 | 3.08 | 1 | |
| 六带线纹鱼Grammistes sexlineatus | 148.7~205.3 | -18.72 | 10.31 | 3.09 | 4 | |
| 冠丝鳍鱼Zu cristatus | 276.7~305.6 | -19.54 | 10.40 | 3.12 | 2 | |
| 横带扁颌针鱼Ablennes hians | 347.6 | -18.07 | 10.68 | 3.20 | 1 | |
| 长鳍舵Kyphosus cinerascens | 144.6~183.4 | -17.65 | 10.88 | 3.26 | 5 | |
| 鲯鳅Coryphaena hippurus | 337.3 | -17.09 | 10.89 | 3.26 | 1 | |
| 符氏眶灯鱼Diaphus fragilis | 74.3~78.1 | -18.45 | 11.12 | 3.33 | 2 | |
| 䲟Echeneis naucrates | 354.3~513.6 | -17.72 | 11.15 | 3.34 | 4 | |
| 瓦氏眶灯鱼Diaphus watasei | 96.3 | -19.51 | 11.30 | 3.38 | 1 | |
| 异鳞蛇鲭Lepidocybium flavobrunneum | 48.9~613.7 | -17.91 | 11.30 | 3.38 | 3 | |
| 黄鳍金枪鱼Thunnus albacares | 1420~1440 | -17.86 | 11.54 | 3.45 | 2 | |
| 灿烂眶灯鱼Diaphus fulgens | 58.3 | -19.07 | 11.59 | 3.47 | 1 | |
| 菲氏眶灯鱼Diaphus phillipis | 47.3~59.2 | -18.53 | 11.61 | 3.47 | 3 | |
| 大鳞魣Sphyraena barracuda | 684.7 | -16.94 | 11.801 | 3.53 | 1 | |
| 头足类 | 鸢乌贼Symplectoteuthis oualaniensis | 164.3~213.3 | -17.60 | 10.10 | 3.03 | 5 |
| 菱鳍乌贼Thysanoteuthis rhombus | 254.6~294.3 | -18.84 | 10.05 | 3.01 | 5 | |
| 紫水孔蛸Tremoctopus gracilis | 98.7~124.7 | -18.48 | 12.33 | 3.69 | 4 | |
| 船蛸Argonauta argo | 137.3 | -17.94 | 12.85 | 3.84 | 1 |
Fig. 2 Stable isotope biplots of δ13C and δ15N values of major fishery species in the central western South China Sea图2 南海中西部海域主要渔业生物的δ13C和δ15N值的双位图 |
Tab. 2 Stable isotope metrics values of main fishery in the central western South China Sea表2 南海中西部海域主要渔业生物稳定同位素参数值 |
| 参数名称 | 简写 | 参数值 |
|---|---|---|
| δ13C 值差 | CR | 3.49 |
| δ15N 值差 | NR | 4.91 |
| 多边形面积 | TA | 9.48 |
| 质心均距 | CD | 1.2 |
| 最短均值 | NND | 1.69 |
| 最短标准差 | SDNND | 0.74 |
Fig. 3 The trophic levels of major fishery species in the central western South China Sea图3 南海中西部海域主要渔业生物的营养级 |
Tab. 3 Different body length (mantle length) of fishery biological trophic level ( TL)表3 不同体长(胴长)渔业生物的营养级 |
| 种 类 | 体长范围/mm | δ13C | δ15N | TL | 尾数 |
|---|---|---|---|---|---|
| 鸢乌贼Symplectoteuthis oualaniensis | 75.6~8.3 | -17.63 | 9.35 | 2.81 | 5 |
| 菱鳍乌贼Thysanoteuthis rhombus | 112.4~132.4 | -18.08 | 9.5 | 2.85 | 5 |
| 164.3~213.3 | -17.6 | 10.09 | 3.03 | 5 | |
| 152.3~202.3 | -18.91 | 10.31 | 3.09 | 2 | |
| 254.6~294.3 | -18.84 | 10.05 | 3.01 | 5 | |
| 红鳍圆鲹Decapterus russelli | 103.3~116.7 | -18.71 | 9.89 | 2.97 | 6 |
| 127.3~134.5 | -18.67 | 10.22 | 3.06 | 3 | |
| 细鳞圆鲹Decapterus lajang | 144.3~224.8 | -17.6 | 9.08 | 2.73 | 14 |
| 257~300 | -16.51 | 9.44 | 2.83 | 6 | |
| 黄鳍金枪鱼Thunnus albacares | 143~149 | -17.38 | 7.63 | 2.3 | 2 |
| 162~167 | -17.65 | 8.29 | 2.5 | 5 | |
| 265~281 | -17.1 | 9.08 | 2.73 | 5 | |
| 1420~1440 | -17.86 | 11.54 | 3.45 | 2 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}