
三亚珊瑚礁珊瑚藻种类及其空间特征与环境因子的关系
|
雷新明(1981—), 男, 河南省潢川县人, 博士, 主要从事珊瑚礁生态学研究。E-mail:leixm@scsio.ac.cn |
Copy editor: 林强
收稿日期: 2018-08-16
要求修回日期: 2018-09-27
网络出版日期: 2019-06-17
基金资助
国家自然科学基金(41306144)
国家自然科学基金(41676150)
国家重点研发计划(2017YFC0506301)
中国科学院战略性先导科技专项(A类)(XDA13020402)
广东省省级科技计划项目(2017B0303014052)
版权
Species composition of coralline algae and its spatial characteristics related to environmental factors in Sanya coral reefs, China
Copy editor: LIN Qiang
Received date: 2018-08-16
Request revised date: 2018-09-27
Online published: 2019-06-17
Supported by
National Natural Science Foundation of China(41306144)
National Natural Science Foundation of China(41676150)
National Key Research and Development Plan(2017YFC0506301)
Strategic Priority Research Program of the Chinese Academy of Sciences(XDA13020402)
Science and Technology Planning Project of Guangdong Province, China(2017B0303014052)
Copyright
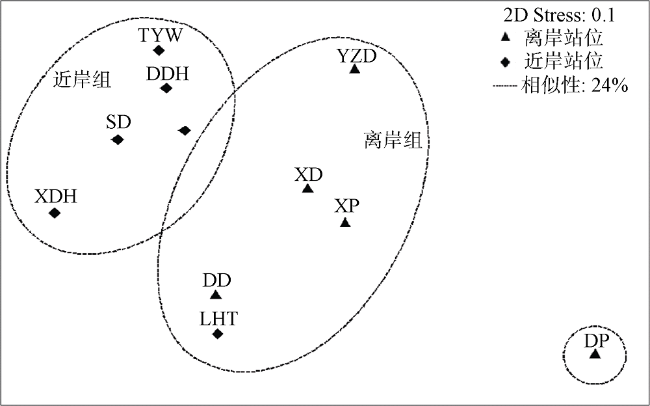
珊瑚藻是珊瑚礁生态系统中重要的功能性群体。2010年通过水肺潜水分析了三亚珊瑚礁区的珊瑚藻种类组成和丰度, 以及群落空间特征与环境因子的关系。共鉴定出3科10属25种珊瑚藻, 其中珊瑚藻科占优势地位。三亚珊瑚礁区的珊瑚藻优势种分别为叉节藻Amphiroa ephedraea、拟中叶藻Mesophyllum simulans、串胞新角石藻Neogoniolithon fosliei、中叶藻M. mesomorphum、圆锥呼叶藻Pneophyllum conicum、孔石藻Porolithon onkodes和布氏水石藻Hydrolithon boergesenii。基于多元统计的分析结果, 将调查采样站位分为近岸型和离岸型2个主要集群, 其中近岸型的珊瑚藻种类以浊度、沉积速率(0~63µm)和悬浮物耐受种类为主, 离岸型则以光依赖-沉积物敏感种类为主。以珊瑚藻种类组成和丰度为基础的群落相似性分析结果表明, 其空间变异处于显著性水平。生物与非生物变量的相关分析结果表明, 最佳两变量组合(浊度和盐度)显著影响三亚珊瑚礁区2种类型调查站位中的珊瑚藻集合。在同一个研究站位, 因受干扰程度不同, 深水处的珊瑚藻的覆盖率比浅水处高。分析结果表明, 在三亚独特的珊瑚礁区中, 自然或人为的物理干扰因素都对调节珊瑚藻群落结构非常重要。
雷新明 , 黄晖 , 练健生 , 张浴阳 , 杨剑辉 . 三亚珊瑚礁珊瑚藻种类及其空间特征与环境因子的关系[J]. 热带海洋学报, 2019 , 38(3) : 79 -88 . DOI: 10.11978/2018084
Coralline algae are important functional group in the coral reef ecosystem. This study was carried out by a series of line transects run underwater with SCUBA apparatus from the reef flat to a depth of 9 m in each of the 11 research stations within the Sanya Coral Reefs in 2010. Among the inshore sites and the offshore sites of Sanya reefs, species composition and abundance of coralline algae were examined in relation to environmental factors. A total of 25 species belonging to three families and 10 genuses were identified, with the family Corallinaceae dominated in the study area. The eight dominant species were Amphiroa ephedraea, Mesophyllum simulans, Neogoniolithon fosliei, M. mesomorphum, Pneophyllum conicum, Porolithon onkodes, and Hydrolithon boergesenii. The sampling stations were divided into inshore type and offshore type based on the results of multivariate statistical analysis. The coralline algae species at the inshore sites were mainly turbidity, deposition rate (0~63 µm) and suspended solids (SS) tolerance species, while the species at the offshore sites were mainly light-dependent and sediment sensitive species. The results of community similarity analysis based on species composition and abundance of coralline algae showed the significant spatial variability. The correlation analysis between biotic and abiotic variables indicated that the best 2-variable combination (turbidity and salinity) showed a significant effect on the pattern of coralline algae assemblages between the two types of research sites in Sanya reefs. The living cover of coralline algae showed that deeper water is higher than that in shallow water due to the different degree of disturbance at the same study sites. This result suggests that physical disturbance, either natural or man-made, is a more important factor in regulating the coralline algae community structure within the coral reef distinct zone of Sanya.
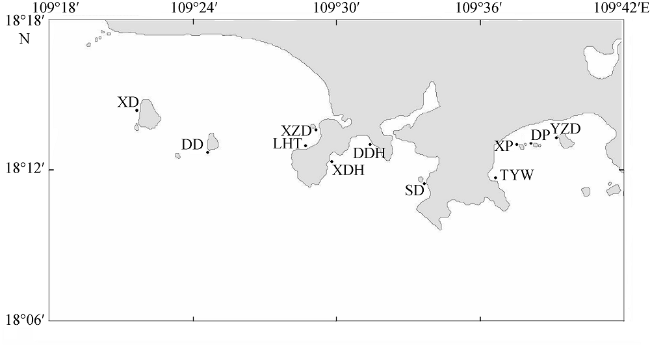
图1 调查站位图XD: 西岛; DD: 东岛; XZH: 小洲岛; LHT: 鹿回头; XDH: 小东海; DDH: 大东海; SD: 神岛; TYW: 太阳湾; XP: 西排; DP: 东排; YZD: 野猪岛 Fig. 1 Study sites in Sanya reefs. XD: Xidao; DD: Dongdao; XZD: Xiaozhoudao; (LHT: Luhuitou; XDH: Xiaodonghai; DDH: Dadonghai; YZD: Yezhudao; DP: Dongpai; XP: Xipai; TYW: Taiyangwan; SD: Shendao |
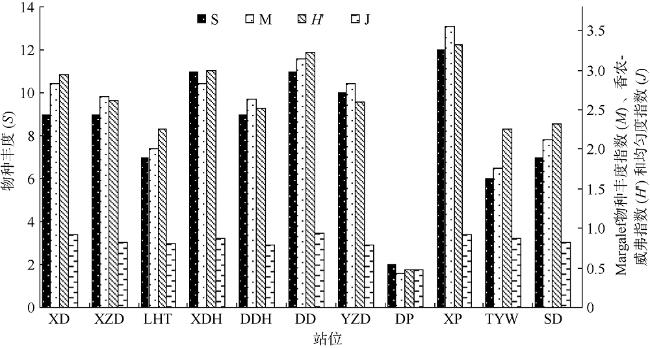
表1 调查区域内的珊瑚藻种类及分布表Tab. 1 Species list and distribution in the study area |
| 珊瑚藻种类 | 站位 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 西岛 | 东岛 | 小洲岛 | 鹿回头 | 小东海 | 大东海 | 野猪岛 | 东排 | 西排 | 太阳湾 | 神岛 | |
| 叉节藻Amphiroa ephedraea | + | + | + | - | + | + | |||||
| 宽扁叉节藻A. anceps | + | + | |||||||||
| 美丽叉节藻 A. beauvoisii | + | ||||||||||
| 矮型石叶藻Lithophyllum pygmaeum | + | + | |||||||||
| *微凹石叶藻L. kotschyanum | + | + | |||||||||
| 三叉新角石藻Neogoniolithon trichotomum | + | ||||||||||
| 太平洋新角石藻N. pacificum | + | + | |||||||||
| 串胞新角石藻N. fosliei | + | + | + | + | + | + | + | + | |||
| 变胞新角石藻N. variabile | + | + | + | + | |||||||
| 锥窝新角石藻N. conicum | + | + | + | + | |||||||
| 巨大新角石藻N. megalocystum | + | ||||||||||
| 撒切尔新角石藻 N. setchellii | + | + | |||||||||
| *小石孔藻Lithoporella melobesioides | + | + | + | + | |||||||
| *太平洋小石孔藻L. pacifica | + | + | + | ||||||||
| *孔石藻Porolithon onkodes | + | + | + | + | |||||||
| *水石藻Hydrolithon reinboldii | + | + | + | + | + | + | + | + | + | ||
| 布氏水石藻H. boergesenii | + | + | + | + | + | + | + | ||||
| 圆锥呼叶藻Pneophyllum conicum | + | + | + | + | |||||||
| 太平洋石枝藻Lithothamnion pacificum | + | + | + | + | |||||||
| 中间石枝藻L. intermedium | + | + | |||||||||
| 尖顶石枝藻L. aculeiferum | + | ||||||||||
| 石枝藻 Lithothamnion sp. | + | ||||||||||
| *中叶藻Mesophyllum mesomorphum | + | + | + | + | + | + | + | + | |||
| *拟中叶藻M. simulans | + | + | + | + | + | + | + | ||||
| *孢石藻Sporolithon erythraeum | + | + | + | + | |||||||
注: *为已有记录的种类 |
表2 各调查站位中3科珊瑚藻分别所占百分比Tab. 2 Contribution of each taxa group (percentage of the total) collected from each study site |
| 站位 | 珊瑚藻所占比例/% | ||
|---|---|---|---|
| 珊瑚藻科 | 混石藻科 | 孢石藻科 | |
| 西岛 | 70.59 | 11.76 | 17.65 |
| 东岛 | 50 | 50 | 0 |
| 小洲岛 | 80.95 | 19.05 | 0 |
| 鹿回头 | 15 | 75 | 10 |
| 小东海 | 67.65 | 32.35 | 0 |
| 大东海 | 95.24 | 0 | 4.76 |
| 神岛 | 72.22 | 27.78 | 0 |
| 太阳湾 | 77.27 | 18.18 | 4.55 |
| 西排 | 77.27 | 18.18 | 4.55 |
| 东排 | 100 | 0 | 0 |
| 野猪岛 | 95.83 | 4.17 | 0 |
表3 各调查站位不同深度的珊瑚藻覆盖率Tab. 3 Living cover of coralline algae of each study site at different depths |
| 站位 | 珊瑚藻覆盖率/% | ||
|---|---|---|---|
| 3m | 6m | 9m | |
| 西岛 | 7.37 | 4.26 | 4.52 |
| 东岛 | 15.76 | 29.64 | 23.09 |
| 小洲岛 | 15.08 | 4.61 | |
| 鹿回头 | 6.97 | 5.06 | |
| 小东海 | 23.27 | 12.73 | 11.61 |
| 大东海 | 15.45 | 8.95 | |
| 神岛 | 2.35 | 0.22 | |
| 太阳湾 | 12.5 | 5.95 | 3.78 |
| 西排 | 38.45 | 27.54 | 7.25 |
| 东排 | 13.91 | 20.32 | 7.40 |
| 野猪岛 | 11.20 | 1.60 | |
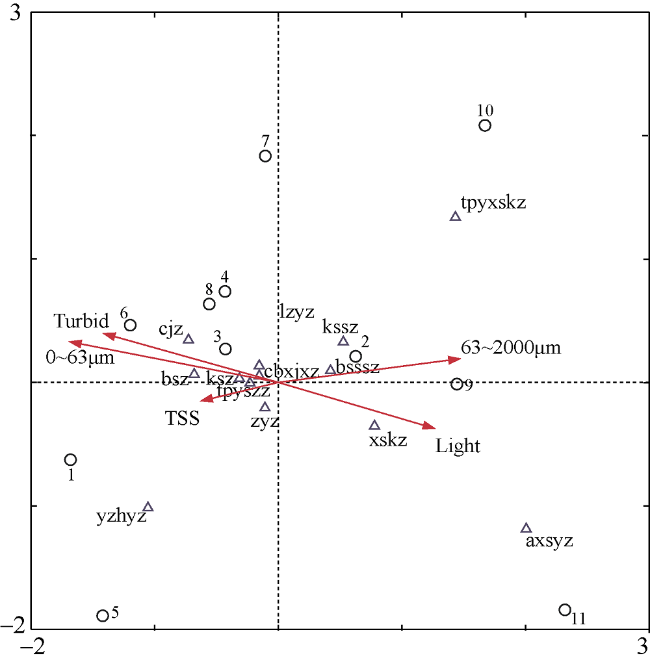
图4 以三亚珊瑚礁区主要珊瑚藻种类组成为响应变量, 以环境因子为解释变量构建CCA排序图Turbid: 浊度; 0~63μm和63~2000μm代表不同粒径的沉积速率; TSS: 总悬浮物含量; Light: 光衰减系数; 数字1~11分别代表站位: 西岛、东岛、小洲岛、鹿回头、小东海、大东海、神岛、太阳湾、西排、东排、野猪岛。cjz: 叉节藻Amphiroa ephedraea; lzyz: 拟中叶藻Mesophyllum simulans; cbxjxz:串胞新角石藻Neogoniolithon fosliei; zyz: 中叶藻Mesophyllum mesomorphum; yzhyz: 圆锥呼叶藻Pneophyllum conicum; axsyz: 矮型石叶藻Lithophyllum pygmaeum; ksz: 孔石藻Porolithon onkodes; tpyxskz: 太平洋小石孔藻Lithoporella pacifica; xskz: 小石孔藻Lithoporella melobesioides; bsssz: 布氏水石藻Hydrolithon boergesenii; kssz: 孔水石藻Hydrolithon onkodes; tpyszz: 太平洋石枝藻Lithothamnion pacificum; bsz: 孢石藻Sporolithon erythraeum Fig. 4 CCA analysis between the main species of coralline algae composition and environmental variables (Turbid: turbidity; 0~63μm and 63~2000μm represent different deposition rates; TSS: total suspended solids; Light: light attenuation coefficient. Numbers from 1 to 11 represent stations: Xidao, Dongdao, Xiaozhoudao, Luhuitou, Xiaodonghai, Dadonghai, Shendao, Taiyangwan, Xipai, Dongpai, Yezhudao. Species abbreviation are as follows. cjz: Amphiroa ephedraea; lzyz: Mesophyllum simulans; cbxjxz: Neogoniolithon fosliei; zyz: Mesophyllum mesomorphum; yzhyz: Pneophyllum conicum; axsyz: Lithophyllum pygmaeum; ksz: Porolithon onkodes; tpyxskz: Lithoporella pacifica; xskz: Lithoporella melobesioides; bsssz: Hydrolithon boergesenii; kssz: Hydrolithon onkodes; tpyszz: Lithothamnion pacificum; bsz: Sporolithon erythraeum |
表4 基于珊瑚藻群落主要种类组成和环境因子之间关系的CCA分析结果Tab. 4 Summary of CCA analysis between the main species of coralline algae composition and environmental variables |
| 轴1 | 轴2 | 总变异 | ||
|---|---|---|---|---|
| 环境因子与排序轴的相关系数 | 浊度 | -0.699 | 0.158 | |
| 光照 | 0.625 | -0.149 | ||
| 总悬浮物含量 | -0.308 | -0.060 | ||
| 沉积速率(0~63µm) | -0.835 | 0.131 | ||
| 沉积速率(63~2000µm) | 0.728 | 0.077 | ||
| 排序轴的 统计结果 | 特征根 | 0.353 | 0.205 | |
| 物种-环境相关性 | 0.935 | 0.835 | ||
| 累计变异百分数 | ||||
| 物种数据 | 20.7 | 32.8 | ||
| 物种-环境数据 | 41.4 | 65.5 | ||
| 特征根之和 | 1.705 | |||
| 典型特征根之和 | 0.853 |
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
李秀保 , 2011. 三亚造礁石珊瑚群落组成、时空分布及主要影响因子识别研究[D]. 北京: 中国科学院研究生院: 1-115.
|
| [2] |
练健生, 黄晖, 黄良民 , 等, 2010. 三亚珊瑚礁及其生物多样性[M]. 北京: 海洋出版社: 1-106.
|
| [3] |
邢帅, 谭烨辉, 周林滨 , 等, 2012. 水体浑浊度对不同造礁石珊瑚种类共生虫黄藻的影响[J]. 科学通报, 57(5):348-354.
|
| [4] |
徐兆礼, 陈亚瞿 , 1989. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系[J]. 生态学杂志, 8(4):13-15.
|
| [5] |
张德瑞, 周锦华 , 1978. 西沙群岛珊瑚藻科的研究Ⅰ[J]. 海洋科学集刊, 12:17-23.
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}