
形态学和SNP标记分析马氏珠母贝杂交子代及其亲本群体的遗传结构
|
黄景(1993—), 男, 湖北省黄石市人, 硕士研究生, 研究方向为马氏珠母贝分子遗传育种。E-mail: |
Copy editor: 林强
收稿日期: 2019-01-21
要求修回日期: 2019-02-27
网络出版日期: 2019-11-26
基金资助
国家贝类产业技术体系建设专项(CARS-49)
版权
Morphological and SNP markers for analysis of genetic structure of hybrid progeny and their parental populations of Pinctada fucata martensii
Copy editor: LIN Qiang
Received date: 2019-01-21
Request revised date: 2019-02-27
Online published: 2019-11-26
Supported by
Modern Agro-industry Technology Research System(CARS-49)
Copyright
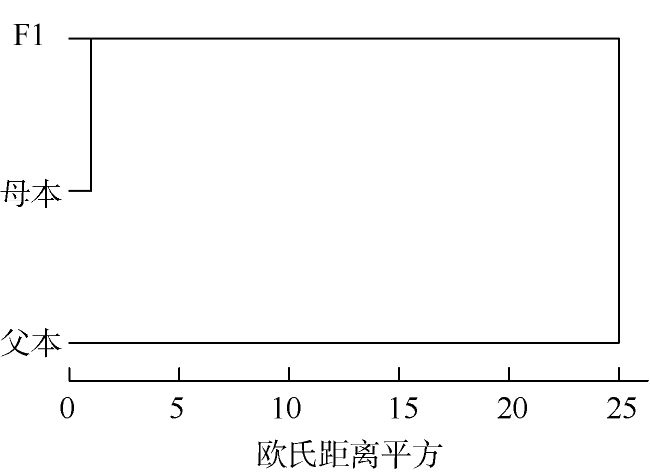
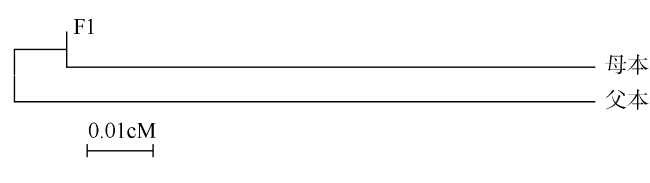
马氏珠母贝是重要的海水养殖贝类, 为了探究繁育亲本及其子代的遗传结构和关系, 本研究运用多元性形态学和SNP标记对马氏珠母贝母本深圳群体、父本海南群体和其杂交子代F1的外部形态和分子遗传结构进行分析。结果发现, 3个群体的综合判别率为72%, F1和母本群体的形态差异最小, 父本群体与母本、F1的形态差异较大。采用HRM法 (high resolution melting) 应用4个SNP位点对这3个群体进行分型, 3个群体的平均观测杂合度Ho和期望杂合度He分别为0.2110~0.2879和0.3317~0.4685, F1的杂合度高于两个亲本; 平均多态信息含量PIC值为0.2643~0.3556, 呈现中等程度遗传多样性。F1与母本之间的基因流Nm最大(7.7701), 遗传距离最小(0.0546), 亲缘关系最近; 两个亲本之间的Nm最小(1.9662), 遗传距离最大(0.1759)。rs8位点可以判别两个亲本群体, 可作为特异性的标记。该结果可以为马氏珠母贝群体遗传结构鉴别、育种群体管理提供指导。
黄景 , 潘肖兰 , 许濛 , 刘文广 , 张华 , 何毛贤 . 形态学和SNP标记分析马氏珠母贝杂交子代及其亲本群体的遗传结构[J]. 热带海洋学报, 2019 , 38(6) : 80 -89 . DOI: 10.11978/2019012
The pearl oyster, Pinctada fucata martensii, is a primary marine bivalve species. To study the genetic structure and relationship of cultured parents and their hybrid generation, external morphology and genetic structure were assessed in the three populations of P. fucata martensii from Shenzhen population (female), Hainan population (male) and their hybrid generation F1, based on morphological multivariate analysis and SNP markers. The results showed that the average discriminant accuracy was 72%, and the morphological characteristics of F1 and female were similar, while male was the most different from the other two populations. Four SNP markers of the three populations were genotyped by the HRM (high resolution melting) method. The results showed that the average observed (Ho) and expected (He) heterozygosity were 0.2110 to 0.2879 and 0.3317 to 0.4685, respectively; and the heterozygosity of F1 was higher than that of the parent populations. The average polymorphism information content (PIC) values of the three populations were in the range of 0.2643 to 0.3556, which showed moderate genetic diversity. Gene flow (Nm) between F1 and female was the largest (7.7701), with minimum genetic distance being 0.0546; thus, their genetic relationships were similar. Gene flow (Nm) between the parent populations was minimum (1.9662), with largest genetic distance being 0.1759. Finally, we found that rs8 marker can be used as a specific marker to discriminate parent populations. These results should assist the discriminate of genetic structure and management of selective breeding of P. fucata martensii.
Key words: Pinctada fucata martensii; genetics; genetic structure; morphology; SNP
壳宽指数 = 壳宽/(壳长+壳高+壳宽)
壳重指数 = [壳重/(壳长×壳高×壳宽)]×105
肥满度指数 = [软体部重/(壳长×壳高×壳宽)]×105
表1 HRM法基因分型引物Tab. 1 Primers used for gene genotyping by HRM |
| 位点名称 | 序列(5'-3') | SNP 类型 | 退火温度/℃ | 对应产物长度/bp |
|---|---|---|---|---|
| rs8 | F: CCCAGCATACTTTATGGCAGAAT R: CCTAGCGTGGAGAAGAGATGTA | T/C | 55 | 122 |
| rs33 | F: CAATACTCTCAGGGACATCTT R: CTCAACAGTGTCGCTTATATG | A/G | 50 | 107 |
| rs35 | F: TTCCGCAGGATGAATTTAGGT R: TCTGAAGTCTGTTTGTCCAATG | C/A | 53 | 103 |
| rs44 | F: CCAAAGCCAACTACACCCAAA R: GTCAAACGCCCACGATACAC | C/A | 55 | 144 |
注: F为正向引物, R为反向引物 |
表2 马氏珠母贝3个群体主要形态参数的平均值Tab. 2 Average values of main morphological parameters in three populations of P. fucata martensii |
| 性状 | 父本 | F1 | 母本 |
|---|---|---|---|
| 壳长 | 58.25±5.58a | 56.40±5.20a | 56.52±4.29a |
| 壳宽 | 23.44±2.23a | 20.23±2.84b | 21.72±2.43ab |
| 壳高 | 59.24±5.10a | 55.60±5.47b | 56.25±4.45b |
| 铰合线 | 51.16±3.79a | 50.17±4.97a | 50.84±4.28a |
| 总重 | 28.16±6.07a | 24.60±7.49b | 24.12±5.33b |
| 壳重 | 14.23±2.78a | 13.11±3.78ab | 11.95±2.62b |
| 软体部重 | 10.77±3.25a | 8.81±3.38b | 8.49±2.64b |
| 壳宽指数 | 0.17±0.01a | 0.15±0.01b | 0.16±0.01ab |
| 壳重指数 | 18.12±2.75b | 20.42±2.06a | 17.43±2.60b |
| 肥满度指数 | 13.36±2.27a | 13.28±2.53a | 13.02±2.25a |
| 壳高/壳长 | 1.02±0.05a | 0.99±0.05b | 1.00±0.06ab |
| 壳宽/壳长 | 0.41±0.03a | 0.36±0.03b | 0.38±0.03ab |
| 铰合线/壳长 | 0.88±0.05b | 0.89±0.05ab | 0.90±0.05a |
注: 上标不同字母之间表示存在显著性差异(P<0.05) |
表3 马氏珠母贝3个群体形态参数主成分分析Tab. 3 Principal component analysis of three populations of P. fucata martensii by morphological parameters |
| 性状 | 负荷值 | ||
|---|---|---|---|
| 主成分1 | 主成分2 | 主成分3 | |
| 总重 | 0.969* | 0.019 | -0.040 |
| 软体部重 | 0.954* | 0.021 | 0.107 |
| 壳高 | 0.937* | -0.052 | -0.030 |
| 壳重 | 0.932* | -0.027 | -0.103 |
| 铰合线长 | 0.901* | 0.064 | 0.069 |
| 壳长 | 0.884* | 0.067 | -0.433 |
| 壳宽指数 | 0.113 | 0.952* | 0.173 |
| 壳宽 | 0.301 | 0.938* | 0.061 |
| 壳宽/壳长 | 0.102 | 0.936* | 0.250 |
| 壳重指数 | 0.184 | -0.758 | 0.088 |
| 肥满度 | 0.454 | -0.531 | 0.393 |
| 铰合线/壳长 | 0.089 | 0.003 | 0.846* |
| 壳高壳/长 | 0.250 | -0.238 | 0.688* |
| 贡献率/% | 43.2 | 27.6 | 12.8 |
注: *表示负荷值大于0.6(表示该因素可以有效的代表其主成分, 主成分贡献率越大表示该主成分越能反映群体之间的形态差异) |
表4 判别函数对马氏珠母贝3个群体观测样本的预测分类及准确率Tab. 4 Predictive classification and accuracy of discriminant for three populations observation samples of P. fucata martensii by discriminant function |
| 群体 | 数量 | 预测分类 | 判别准确率 | 综合判别率 | |||
|---|---|---|---|---|---|---|---|
| 父本 | F1 | 母本 | P1 | P2 | |||
| 父本 | 50 | 36 | 9 | 5 | 72% | 75% | 72% |
| F1 | 50 | 8 | 32 | 10 | 64% | 68.1% | |
| 母本 | 50 | 4 | 6 | 40 | 80% | 72.7% | |
注: 用逐步判别法建立3个群体的判别函数, 然后在此基础上对所有个体的类别进行判别, 计算判别准确率的公式为: 判别准确率P1 = (判别正确的贝数/实测贝数)×100%; 判别准确率P2=(判别正确的贝数/判别贝数)×100%; 综合判别率= |
表5 马氏珠母贝3个群体遗传多样性参数Tab. 5 Genetic diversity parameters of three populations of P. fucata martensii |
| 群体名称 | 位点 | I | Ho | He | PIC | HWE P值 |
|---|---|---|---|---|---|---|
| 父本 | rs8 | 0.3768 | 0.2500 | 0.2222 | 0.1948 | 0.4527 |
| rs33 | 0.6809 | 0.3438 | 0.4955 | 0.3685 | 0.0780 | |
| rs35 | 0.3111 | 0.0625 | 0.1726 | 0.1555 | 0.0044* | |
| rs44 | 0.6211 | 0.1875 | 0.4365 | 0.3384 | 0.0010* | |
| 平均值 | 0.4975 | 0.2110 | 0.3317 | 0.2643 | ||
| F1 | rs8 | 0.6002 | 0.1515 | 0.4163 | 0.3260 | 0.0003* |
| rs33 | 0.6816 | 0.3030 | 0.4960 | 0.3692 | 0.0230 | |
| rs35 | 0.5860 | 0.4242 | 0.4028 | 0.3182 | 0.7500 | |
| rs44 | 0.6635 | 0.2727 | 0.4779 | 0.3598 | 0.0120 | |
| 平均值 | 0.6328 | 0.2879 | 0.4483 | 0.3433 | ||
| 母本 | rs8 | 0.6881 | 0.4333 | 0.5034 | 0.3724 | 0.4380 |
| rs33 | 0.5799 | 0.3333 | 0.3977 | 0.3137 | 0.3622 | |
| rs35 | 0.6657 | 0.0333 | 0.4808 | 0.3614 | 0.0000* | |
| rs44 | 0.6926 | 0.2333 | 0.4921 | 0.3747 | 0.0025* | |
| 平均值 | 0.6566 | 0.2583 | 0.4685 | 0.3556 |
注: I为香农多样性指数, Ho为观测杂合度, He为期望杂合度, PIC为多态信息含量, HWE为哈迪-温伯格平衡检验; *表示经过校正后仍显著偏离哈迪-温伯格平衡的位点(P<0.01) |
表6 马氏珠母贝3个群体SNP的基因型和频率Tab. 6 Genotypes and frequencies of SNP in three populations of P. fucata martensii |
| 群体名称 | 基因型(频率) | |||
|---|---|---|---|---|
| rs8 | rs33 | rs35 | rs44 | |
| 父本 | TT(0.75) | GG(0.41) | CC(0.88) | CC(0.59) |
| TC(0.25) | GA(0.34) | CA(0.06) | CA(0.19) | |
| CC(0) | AA(0.25) | AA(0.06) | AA(0.22) | |
| F1 | TT(0.64) | GG(0.27) | CC(0.52) | CC(0.24) |
| TC(0.15) | GA(0.30) | CA(0.42) | CA(0.27) | |
| CC(0.21) | AA(0.42) | AA(0.06) | AA(0.49) | |
| 母本 | TT(0.23) | GG(0.10) | CC(0.60) | CC(0.37) |
| TC(0.43) | GA(0.33) | CA(0.03) | CA(0.23) | |
| CC(0.33) | AA(0.57) | AA(0.37) | AA(0.40) | |
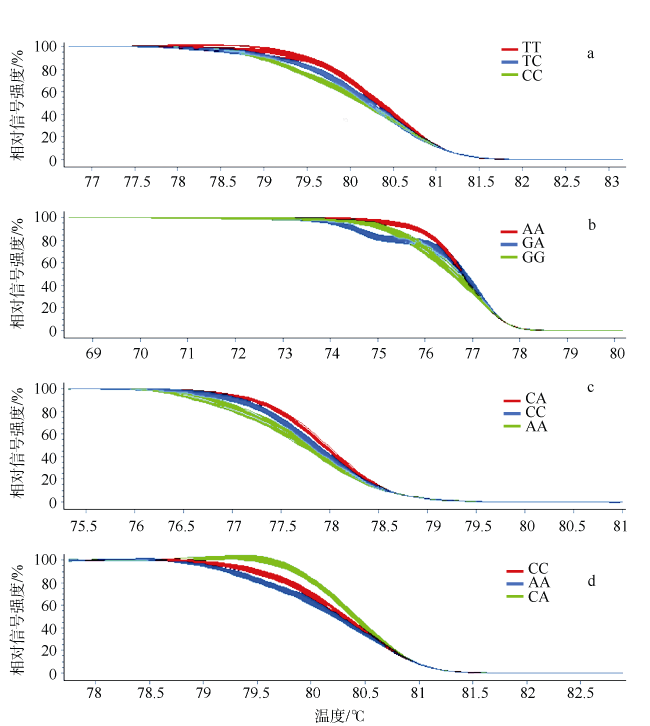
图2 马氏珠母贝3个群体4个位点分型结果图a、b、c、d分别为rs8、rs33、rs35、rs44的HRM分型结果图, 不同颜色代表不同基因型 Fig. 2 The results of four loci typing in tree populations of P. fucata martensii. a, b, c and d are the results of HRM typing for rs8, rs33, rs35, and rs44, respectively, different colours represent different genotypes |
表7 马氏珠母贝3个群体遗传结构参数Tab. 7 Genetic structural parameters of three populations of P. fucata martensii |
| 遗传距离和遗传相似系数 | 基因流和遗传分化系数 | ||||||
|---|---|---|---|---|---|---|---|
| 父本 | F1 | 母本 | 父本 | F1 | 母本 | ||
| 父本 | **** | 0.9320 | 0.8387 | **** | 0.0546 | 0.1128 | |
| F1 | 0.0704 | **** | 0.9947 | 4.3262 | **** | 0.0166 | |
| 母本 | 0.1759 | 0.0546 | **** | 1.9662 | 7.7701 | **** | |
注: 遗传距离和遗传相似系数中, ****下方数据为Nei's遗传距离, ****上方数据为遗传相似系数; 基因流和遗传分化系数中, ****下方数据为基因流Nm值, ****上方数据为遗传分化系数。基因流Nm值 = 0.25(1- Fst) / Fst, Fst为遗传分化系数 |
| 1 |
白牡丹, 王彩虹, 殷豪 , 等, 2012. 苹果不同HRM反应体系分析效果评价[J]. 分子植物育种, 10(1):115-120.
|
| 2 |
陈亮, 赵柏淞, 李玲 , 等, 2014. 人工诱导栉孔扇贝雌核发育胚胎的SNP标记分析[J]. 中国海洋大学学报, 44(2):48-52.
|
| 3 |
陈蓉, 刘建勇, 唐连俊 , 等, 2009. 中国5个海区不同群体毛蚶形态差异分析[J]. 海洋科学, 33(7):64-69.
|
| 4 |
董志国, 李家乐, 郑汉丰 , 2008. 三角帆蚌3个地理种群自交与杂交F1代的形态差异分析[J]. 大连水产学院学报, 23(2):92-97.
|
| 5 |
杜晓东, 李广丽, 刘志刚 , 等, 2002. 合浦珠母贝2个野生种群的遗传多样性[J]. 中国水产科学, 9(2):100-105.
|
| 6 |
谷龙春, 李金碧, 喻达辉 , 等, 2010. 合浦珠母贝双列杂交家系的建立与遗传分析[J]. 水产学报, 34(1):26-31.
|
| 7 |
顾志峰, 王嫣, 石耀华 , 等, 2009. 马氏珠母贝两个不同地理种群的形态性状和贝壳珍珠质颜色比较分析[J]. 渔业科学进展, 30(1):79-86.
|
| 8 |
侯战辉, 王嫣, 石耀华 , 等, 2008. 马氏珠母贝(Pinctada martensii)2个不同地理种群遗传变异的EST-SSR分析[J]. 海洋与湖沼, 39(2):178-183.
|
| 9 |
胡笑蓉, 毛雄英, 王卫华 , 等, 2012. 一种基于HRM技术的快速DNA甲基化检测方法[J]. 中国卫生检验杂志, 22(6):1336-1338.
|
| 10 |
姜因萍, 何毛贤, 林岳光 , 2007. 马氏珠母贝群体内遗传多样性的ISSR分析[J]. 海洋通报, 26(5):62-66.
|
| 11 |
金玉琳 , 2014. 长牡蛎SNP标记开发及其在家系分析和物种鉴定中的应用[D]. 青岛: 中国海洋大学: 32-38.
|
| 12 |
李纪勤 , 2012. 栉孔扇贝(Chlamys farreri)EST-SNP的开发及其应用[D]. 青岛: 中国海洋大学: 53-54.
|
| 13 |
李梅, 吕伟朋, 韦鸿 , 等, 2012. HRM方法检测非小细胞肺癌患者胸水标本癌细胞基因突变的临床意义[J]. 大连医科大学学报, 34(4):321-323, 347.
|
| 14 |
李太武, 孙修勤, 刘艳 , 等, 2002. 中日栉孔扇贝杂交子一代群体的遗传变异[J]. 高技术通讯, 12(6):101-105, 100.
|
| 15 |
李耀国, 刘文广, 林坚士 , 等, 2016. 马氏珠母贝SNP标记开发及家系遗传多态性分析[J]. 海洋通报, 35(1):96-102.
|
| 16 |
刘泽浩, 李捷, 杨学玲 , 等, 2010 中国北方沿海潮间带常见镜蛤的种类[J]. 海洋科学, 34(12):30-35.
|
| 17 |
罗会, 刘宝锁, 黎火金 , 等, 2013. 合浦珠母贝不同地理种群的形态差异和判别分析[J]. 广东农业科学, 40(12):171-174.
|
| 18 |
吕林兰, 杜晓东, 王嫣 , 等, 2008. 马氏珠母贝3个野生种群及种群间杂交后代遗传多样性的ISSR分析[J]. 水生生物学报, 32(1):26-32.
|
| 19 |
曲艳波, 叶翚, 石耀华 , 等, 2006. 马氏珠母贝杂交选育群体遗传变异的RAPD分析[C]// 海南生物技术研究与发展研讨会论文集. 三亚: 中国生物工程学会: 27-34.
|
| 20 |
沈恩健 , 2010. 利用AFLP和SSR对南海区域马氏珠母贝群体的遗传结构分析[D]. 湛江: 广东海洋大学.
|
| 21 |
苏天凤, 蔡云川, 张殿昌 , 等, 2002. 合浦珠母贝3个养殖群体的RAPD分析[J]. 中国水产科学, 9(2):106-109.
|
| 22 |
汤健, 管云雁, 刘文广 , 等, 2013. 马氏珠母贝家系遗传结构的微卫星分析[J]. 海洋科学, 37(8):35-41.
|
| 23 |
童晓飞, 郭莹, 丛晓霏 , 2012. 高分辨溶解曲线及其在检测海洋经济贝类SNPs的应用前景[J]. 北京农业, ( 15):139.
|
| 24 |
王爱民, 石耀华 , 2007. 马氏珠母贝的遗传改良技术[M] //王清印. 海水养殖生物的细胞工程育种. 北京: 海洋出版社: 133-164.
|
| 25 |
王爱民, 王嫣, 顾志峰 , 等, 2010. 马氏珠母贝(Pinctada martensii)2个地理群体杂交子代的杂种优势和遗传变异[J]. 海洋与湖沼, 41(1):140-147.
|
| 26 |
王家丰 , 2013. 长牡蛎基因区SNP标记规模开发及其在遗传育种研究中的应用[D]. 青岛: 中国科学院研究生院(海洋研究所).
|
| 27 |
王艳, 赵雪, 姜振峰 , 等, 2015. 大豆小片段法HRM基因分型体系优化[J]. 中国油料作物学报, 37(4):453-461.
|
| 28 |
杨润婷, 吴波, 李翀 , 等, 2013. 两种SNP分型方法的比较及其在柚品种鉴定中的应用[J]. 园艺学报, 40(6):1061-1070.
|
| 29 |
喻达辉, 李有宁, 吴开畅 . 2005. 中国、日本和澳大利亚珍珠贝的ITS2序列特征分析[J]. 南方水产, ( 02):4-9.
|
| 30 |
张莉 , 2007. 中国珍珠产业的问题、困境与出路[J]. 农业现代化研究, 28(4):443-445.
|
| 31 |
赵杰, 游新勇, 徐贞贞 , 等, 2018. SNP检测方法在动物研究中的应用[J]. 农业工程学报, 34(4):299-305.
|
| 32 |
赵琼一, 李信, 周德贵 , 等, 2010. 后基因组时代下作物的SNP分型方法[J]. 分子植物育种, 8(1):125-133.
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|
| 55 |
|
| 56 |
|
| 57 |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}