
Journal of Tropical Oceanography >
Effects of breeding density on the growth, metabolic enzyme activity and related gene expression level of juvenile Pampus argenteus
Received date: 2019-05-07
Request revised date: 2019-07-25
Online published: 2020-03-10
Supported by
National Natural Science Foundation of China(31772869)
National Natural Science Foundation of China(31872586)
Natural Science Foundation of Zhejiang Province(LY15C190003)
Natural Science Foundation of Zhejiang Province(LY18C190008)
Special Funds for Key Project of Science and Technology of Ningbo(2019C02059)
Copyright
The feeding experiment lasted for 40 days to explore the effects of breeding density (30, 60 and 90 individuals·m-3) on the growth, enzyme activity and mRNA expression of aspartate aminotransferase (AST), alanine aminotransferase (ALT) and lactate dehydrogenase (LDH) in liver and kidney of juvenile Pampus argenteus with an average body weight of (3.88±0.72) g. Results are as follows. The weight gain rate and specific growth rate of the 60 individuals·m -3 group were (235.19±10.23)% and (4.03±0.10)%, respectively, which were significantly higher than those in the 30 individuals·m-3 group (p<0.05), but showed no significant differences with the 90 individuals·m-3 group (p>0.05). The changes of enzyme activity of juvenile Pampus argenteus were the most significant in the 90 individuals·m-3 group and were the least significant in the 30 individuals·m-3group. The activities of LDH, ALT and AST in liver and kidney increased first and then decreased, and the values of the three enzyme activities reached the peak at 6 h, 10 d and 10 d, respectively, which were 2.0 times, 4.3 times and 2.0 times of the initial values. The peak values of the three enzyme activities appeared at 1 d, 3 d and 5 d in kidney, respectively, and the values were 2.1 times, 2.2 times and 3.1 times of the initial values. The changes of mRNA expression of AST, ALT and LDH genes and the changes of the three enzyme activities in liver and kidney showed significant correlations. In conclusion, the breeding density of 60 individuals·m-3 could promote the growth of juvenile Pampus argenteus. However, the 90 individuals·m-3 group might cause extra energy demand increases in Pampus argenteus juveniles. Increased expression of gluconeogenesis-related genes mRNA resulted in significant increases of AST, ALT and LDH activities in liver and kidney, and eventually slowed down the growth of juvenile Pampus argenteus.
NI Jiahao , ZHU Xiaojing , JI Yiping , ZHOU Bin , WANG Yajun , XU Shanliang , WANG Danli . Effects of breeding density on the growth, metabolic enzyme activity and related gene expression level of juvenile Pampus argenteus[J]. Journal of Tropical Oceanography, 2020 , 39(2) : 54 -64 . DOI: 10.11978/2019045
表1 AST、ALT和LDH基因荧光定量PCR所用引物Tab. 1 Primer used for AST, ALT and LDH qRT-PCR |
| 引物 | 序列(5'—3') | 片段大小/bp | 退火温度/℃ |
|---|---|---|---|
| AST-F AST-R | AGTGGAAGGAGATTGCTGACATTGT ACTCCACTCTCTTTGCCTCATCTGT | 242 | 58 |
| ALT-F ALT-R | TCATCAAACTGGTGTCGGTGCG TGTGGTGTAGGAGGGCTCATCAGGC | 103 | 61 |
| LDH-F LDH-R | GCGACAAAGACTACAGCGTGAC CGATGTTGGGGATGATAAACTTGAA | 137 | 57 |
表2 养殖密度对银鲳幼鱼体生长的影响Tab. 2 Effect of density on the growth of juvenile Pampus argenteus |
| 组别 | 初始体质量/g | 终末体质量/g | 增重率/% | 特定生长率/% |
|---|---|---|---|---|
| 低密度组(30尾·m-3) | 4.30±0.50 | 11.27±0.46 | 162.02±10.80 b | 3.21±0.14 b |
| 中密度组(60尾·m-3) | 3.60±0.30 | 12.07±0.37 | 235.19±10.23 a | 4.03±0.10 a |
| 高密度组(90尾·m-3) | 3.75±0.45 | 11.10±0.91 | 196.00±24.25 ab | 3.61±0.27 ab |
注: 不同小写字母表示该指标在不同密度组中差异显著(p<0.05) |
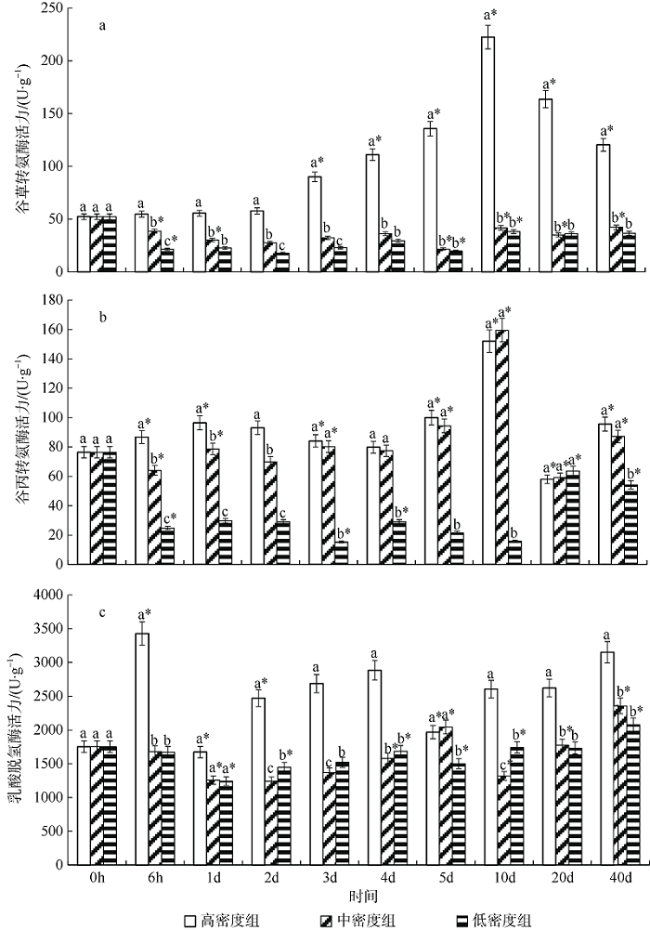
图1 养殖密度对银鲳幼鱼肝脏谷草转氨酶(a)、谷丙转氨酶(b)、乳酸脱氢酶(c)活力的影响*表示同一密度组与前一时间点比较有显著差异(p<0.05); 不同小写字母表示不同密度组在同一时间点差异显著(p<0.05) Fig. 1 Effect of density on liver AST (a), ALT (b) and LDH (c) activity of juvenile Pampus argenteus. * indicates significant difference between different treatment times at the same density (p<0.05), Different small letters above the bars indicate significant differences between different densities at the same time (p<0.05) |
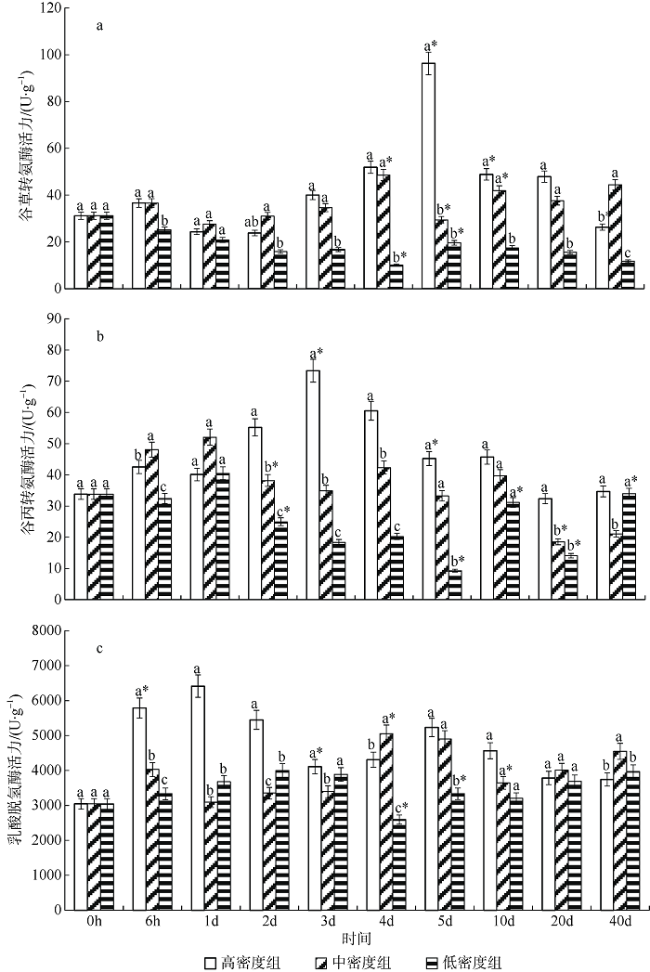
图2 养殖密度对银鲳幼鱼肾脏谷草转氨酶(a)、谷丙转氨酶(b)、乳酸脱氢酶(c)活力的影响*表示同一密度组与前一时间点比较有显著差异(p<0.05); 不同小写字母表示不同密度组在同一时间点差异显著(p<0.05) Fig. 2 Effect of density on kidney AST (a), ALT (b) and LDH (c) activity of juvenile Pampus argenteus. Asterisk (*) indicates significant difference between different treatment times at the same density (p<0.05), Different small letters above the bars indicate significant differences between different densities at the same time (p<0.05) |
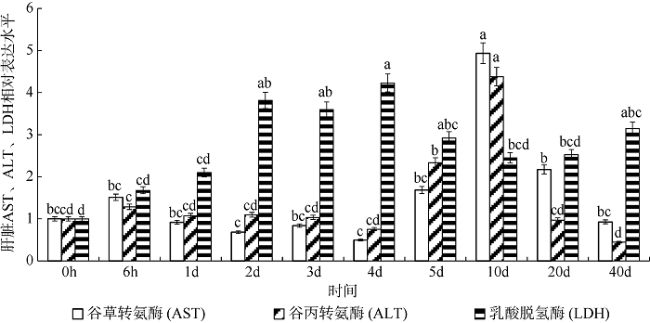
图3 高密度组(90尾·m-3)银鲳幼鱼肝脏AST、ALT、LDH 3种酶基因的mRNA表达变化不同小写字母表示该种酶在不同时间点差异显著(p<0.05) Fig. 3 Change of mRNA relative expression on three liver enzymes in high density group of juvenile Pampus argenteus. Different small letters above the bars indicate significant differences between different times at the same enzyme (p<0.05) |
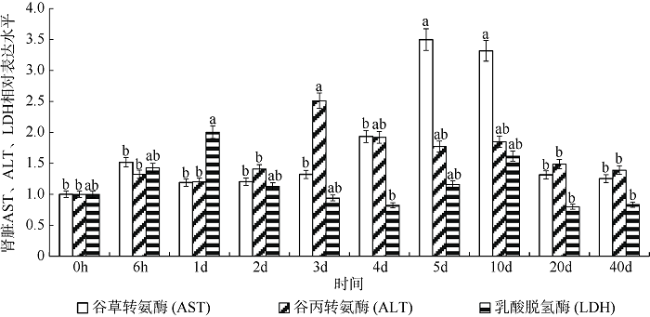
图4 高密度组(90尾·m-3)银鲳幼鱼肾脏AST、ALT、LDH 3种酶基因的mRNA表达变化不同小写字母表示该种酶在不同时间点差异显著(p<0.05) Fig. 4 Change of mRNA relative expression on three kidney enzymes in high density group of juvenile Pampus argenteus. Different small letters above the bars indicate significant differences between different times at the same enzyme (p<0.05). |
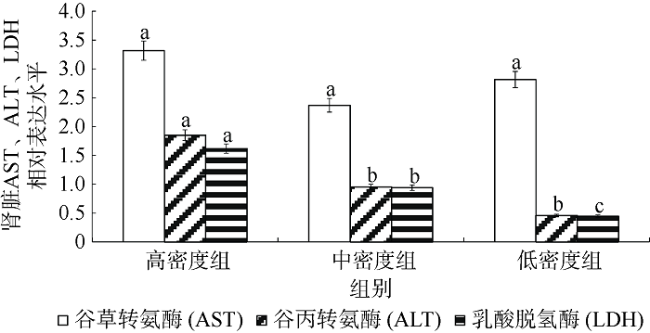
图5 不同密度下银鲳幼鱼肝脏AST、ALT、LDH 3种酶基因的mRNA表达差异不同小写字母表示该种酶在不同密度下差异显著(p<0.05) Fig. 5 Difference of mRNA relative expression on three liver enzymes between different density stresses of juvenile Pampus argenteus. Different small letters above the bars indicates significant differences between different densities at the same enzyme (p<0.05) |
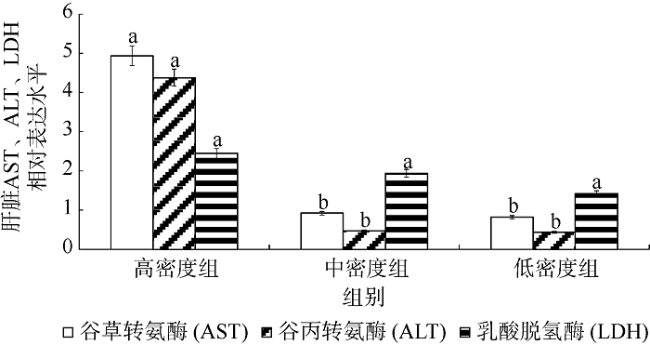
图6 不同密度下银鲳幼鱼肾脏AST、ALT、LDH 3种酶基因的mRNA表达差异不同小写字母表示该种酶在不同密度组差异显著(p<0.05) Fig. 6 Difference of mRNA relative expression on three kidney enzymes between different density stresses of juvenile Pampus argenteus. Different small letters above the bars indicate significant differences between different densities at the same enzyme (p<0.05) |
| 1 |
李爱华 , 1997. 拥挤胁迫对草鱼血浆皮质醇、血糖及肝脏中抗坏血酸含量的影响[J]. 水生生物学报, 21(4):384-386.
LI AIHUA, 1997. Effects of crowding stress on plasma cortisol, glucose and liver ascorbic acid in grass carp(Ctenopharyngodon idellus)[J]. Acta Hydrobiologica Sinica, 21(4):384-386 (in Chinese with English abstract).
|
| 2 |
李大鹏, 庄平, 严安生 , 等, 2004. 光照、水流和养殖密度对史氏鲟稚鱼摄食、行为和生长的影响[J]. 水产学报, 28(1):54-61.
LI DAPENG, ZHUANG PING, YAN ANSHENG, et al, 2004. The influences of illumination, water current and stocking density on feeding, behavior and growth in juveniles Acipenser schrenckii[J]. Journal of Fisheries of China, 28(1):54-61 (in Chinese with English abstract).
|
| 3 |
梁从飞 , 2015. 尼罗罗非鱼耐盐碱选育一代耐受性、生长性能及鳃Na+/HCO3-共转运子、碳酸酐酶基因表达[D]. 硕士学位论文, 上海:上海海洋大学.
LIANG CONGFEI, 2015. Tolerance, growth and branchial mRNA expression of Na+/HCO3- cotransporter and CA of the first selected generation of Oreochromis niloticus in saline and alkaline water[D]. Shanghai: Shanghai Ocean University (in Chinese with English abstract).
|
| 4 |
逯尚尉 , 2010. 不同饵料、密度和光周期对点带石斑鱼(Epinephelus malabaricus)幼鱼的影响[D] . 硕士学位论文, 南京: 南京农业大学.
LU SHANGWEI, 2010. Effect of different diets, stocking densities and photoperiods on juvenile Epinephelus malabaricus[D] . Nanjing: Nanjing Agricultural University (in Chinese with English abstract).
|
| 5 |
逯尚尉, 刘兆普, 余燕 , 2011. 密度胁迫对点带石斑鱼幼鱼生长、代谢的影响[J]. 中国水产科学, 18(2):322-328.
LU SHANGWEI, LIU ZHAOPU, YU YAN, 2011. Effects of density stress on growth and metabolism of juvenile Epinephelus malabaricus[J]. Journal of Fishery Sciences of China, 18(2):322-328 (in Chinese with English abstract).
|
| 6 |
彭士明, 施兆鸿, 孙鹏 , 等, 2010. 养殖密度对银鲳幼鱼生长及组织生化指标的影响[J]. 生态学杂志, 29(7):1371-1376.
PENG SHIMING, SHI ZHAOHONG, SUN PENG, et al, 2010. Effects of breeding density on the growth and tissues biochemical indices of juvenile silver pomfret(Pampus argenteus)[J]. Chinese Journal of Ecology, 29(7):1371-1376 (in Chinese with English abstract).
|
| 7 |
亓成龙 , 2016. 养殖密度和亚硝酸盐胁迫对团头鲂幼鱼生理机能和肌肉品质的影响[D]. 硕士学位论文, 武汉: 华中农业大学.
QI CHENGLONG, 2016. Effect of stocking density and nitrite on physiological functions and muscle quality of blunt snout bream Megalobrama amblycephala juveniles[D] . Wuhan: Huazhong Agricultural University (in Chinese with English abstract).
|
| 8 |
强俊, 徐跑, 何杰 , 等, 2011. 氨氮与拥挤胁迫对吉富品系尼罗罗非鱼幼鱼生长和肝脏抗氧化指标的联合影响[J]. 水产学报, 35(12):1837-1848.
QIANG JUN, XU PAO, HE JIE, et al, 2011. The combined effects of external ammonia and crowding stress on growth and biochemical activities in liver of (GIFT) Nile tilapia juvenile(Oreochromis niloticus)[J]. Journal of Fisheries of China, 35(12):1837-1848 (in Chinese with English abstract).
|
| 9 |
强俊, 杨弘, 何杰 , 等, 2014. 3种品系尼罗罗非鱼生长及高密度胁迫后生理响应变化的比较[J]. 中国水产科学, 21(1):142-152.
QIANG JUN, YANG HONG, HE JIE, et al, 2014. Comparison on growth performance in three different strains of Nile tilapia and physiological responses after short-term high stocking density stress[J]. Journal of Fishery Sciences of China, 21(1):142-152 (in Chinese with English abstract).
|
| 10 |
任源远 , 2014. 养殖密度对施氏鲟大规格鱼种生长性能及血液生理生化组分的影响[D]. 硕士学位论文, 山东: 中国海洋大学.
REN YUANYUAN, 2014. Effects of stocking density on growth performance and blood physiological and biochemical components in large-sized juvenile Amur sturgeon[D]. Shandong: Ocean University of China (in Chinese with English abstract).
|
| 11 |
石小涛 , 2006. 史氏鲟在拥挤胁迫中的应激反应[D]. 硕士学位论文, 华中农业大学, 2006.
SHI XIAOTAO, 2006. The stress-influences of rearing densities on juvenile Amur sturgeon, Acipenser schrenckii[D]. Wuhan: Huazhong Agricultural University (in Chinese with English abstract).
|
| 12 |
孙大川, 谭洪新, 罗国芝 , 等, 2009. 养殖密度对宝石鲈生长性能和血液生化指标的影响[J]. 渔业现代化, 36(2):12-15.
SUN DACHUAN, TAN HONGXIN, LUO GUOZHI, et al, 2009. Effect of stocking density on growth and blood biochemical parameters of Scortum barcoo[J]. Fishery Modernization, 36(2):12-15 (in Chinese with English abstract).
|
| 13 |
王春枝, 李忠, 梁宏伟 , 等, 2014. 低氧胁迫对鲢线粒体ATP酶活性及F1-δ基因表达的影响[J]. 中国水产科学, 21(3):454-463.
WANG CHUZHI, LI ZHONG, LIANG HONGWEI, et al, 2014. The effects of hypoxia stress on mitochondrial ATPase activity and the expression of F1-δ in Hypophthalmichthys molitrix[J]. Journal of Fishery Sciences of China, 21(3):454-463 (in Chinese with English abstract).
|
| 14 |
王文博, 汪建国, 李爱华 , 等, 2004. 拥挤胁迫后鲫鱼血液皮质醇和溶菌酶水平的变化及对病原的敏感性[J]. 中国水产科学, 11(5):408-412.
WANG WENBO, WANG JIANGUO, LI AIHUA, et al, 2004. Changes of cortisol and lysozyme levels in Carassius auratus blood after crowding stress and the fish sensitivity to pathogen[J]. Journal of Fishery Sciences of China, 11(5):408-412 (in Chinese with English abstract).
|
| 15 |
王跃斌, 胡成硕, 胡则辉 , 等, 2017. 养殖密度对日本黄姑鱼幼鱼生长及非特异性免疫的影响[J]. 海洋渔业, 39(1):51-57 .
WANG YUEBIN, HU CHENGSHUO, HU ZEHUI, et al, 2017. Effects of different stocking densities on growth performance and non-specific immunity of juvenile Nibea japonica[J]. Marine Fisheries, 39(1):51-57 (in Chinese with English abstract).
|
| 16 |
夏华, 李威名, 陈阿琴 , 等, 2013. 两种养殖密度下鲫鱼生长和免疫酶活性的比较[J]. 广东农业科学, 40(12):130-133.
XIA HUA, LI WEIMING, CHENG AQIN, et al, 2013. Comarative analysis of growth and immune enzyme activities of Carassius auratus under two densities[J]. Guangdong Agricultural Sciences, 40(12):130-133 (in Chinese with English abstract)
|
| 17 |
薛宝贵, 楼宝, 徐冬冬 , 等, 2013. 密度胁迫对黄姑鱼幼鱼生长、代谢及非特异性免疫的影响[J]. 渔业科学进展, 34(2):45-51.
XUE BAOGUI, LOU BAO, XU DONGDONG, et al, 2013. Impact of density stress on growth, metabolism and non-specific immune functions of juvenile Nibea albiflora[J]. Progress in fishery sciences, 34(2):45-51 (in Chinese with English abstract).
|
| 18 |
张建明, 郭柏福, 高勇 , 2013. 中华鲟幼鱼对慢性拥挤胁迫的生长、摄食及行为反应[J]. 中国水产科学, 20(3):592-598.
ZHANG JIANMING, GUO BOFU, GAO YONG, 2013. Effects of chronic crowding stress on growth, feeding, and behavior in Chinese sturgeon Acipenser sinensis F2 juveniles[J]. Journal of Fishery Sciences of China, 20(3):592-598 (in Chinese with English abstract).
|
| 19 |
于淼, 樊启学, 程鹏 , 等, 2008. 急性拥挤胁迫对鲤血液中皮质醇及几项生化指标的影响[J]. 淡水渔业, 38(4):20-24 .
YU MIAO, FAN QIXUE, CHENG PENG, et al, 2008. Effects of acute crowding stress on cortisol and several biochemical indexes in Cyprinus carpio serum[J]. Freshwater Fisheries, 38(4):20-24 (in Chinese with English abstract).
|
| 20 |
张磊, 樊启学, 赵志刚 , 等, 2007. 慢性拥挤胁迫对鲤生长及血液生化指标的影响[J]. 大连海洋大学学报, 22(6):465-469.
ZHANG LEI, FAN QIXUE, ZHAO ZHIGANG, et al, 2007. The effects of chronic crowding stress on growth and blood Biochemical indexes in common carp Cyprinus carpio[J]. Journal of Dalian Fisheries University, 22(6):465-469 (in Chinese with English abstract).
|
| 21 |
张廷军, 杨振才, 孙儒泳 , 1998. 鱼类对高密度环境的适应[J]. 水产科技情报, 25(3):110-113.
ZHANG TINGJUN, YANG ZHENCAI, SUN RUYONG, 1998. The Acclimation of Fishes to High Stocking Density[J]. Fisheries Science & Technology Information, 25(3):110-113 (in Chinese with English abstract).
|
| 22 |
张曦文, 吴垠, 贺茹靖 , 等, 2012. 循环水养殖模式下养殖密度对青石斑鱼生长及生理指标的影响[J]. 大连海洋大学学报, 27(6):518-522.
ZHANG XI WEN, WU YIN, HE RUJING, et al, 2012. The effects of stocking density on growth and physiological indices of grouper Epinephelus awoara in recirculating aquaculture[J]. Journal of Dalian Ocean University, 27(6):518-522 (in Chinese with English abstract).
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
Hoener, G,
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}