
Journal of Tropical Oceanography >
Growth characteristics of four representative fishes and their responses to seagrass resource changes in typical tropical seagrass beds of Hainan Island
Copy editor: YIN Bo
Received date: 2020-01-12
Request revised date: 2020-03-09
Online published: 2020-03-19
Supported by
National Natural Science Foundation of China(41730529)
National Natural Science Foundation of China(41806147)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0405)
Guangzhou Science and Technology Program(201904010370)
Innovation Academy of South China Sea Ecology and Environmental Engineering, Chinese Academy of Sciences(ISEE2018PY01)
Innovation Academy of South China Sea Ecology and Environmental Engineering, Chinese Academy of Sciences(ISEE2018ZD02)
Copyright
The functions of seagrass beds to provide food and shelter in response to changes in seagrass coverage are still unclear. Xincun Bay, Li’an Port and Tanmen Port seagrass beds of Hainan Island were chosen as our research areas; and Siganus fuscessens, Siganus guttatus, Pelates quadrilineatus, and Terapon jarbua were selected for our research. The relationships between body length and body mass of these four representative fishes, and their responses to seagrass resource changes were addressed. The results showed that the four species inhabiting the Tanmen seagrass bed had negative allometric growth, and the ones in other seagrass beds had positive allometric or constant growth. The body mass growth rate (allometric growth factor b) and the fatness of these four fishes showed the trend of Li’an Port > Xincun Bay > Tanmen Port, which was consistent with the trend of seagrass coverage, density and biomass of the three seagrass beds. We speculate that the decreasing resources of seagrass increase the predation pressure of fish and reduce their food sources, and might consequently decrease growth rates of these representative fishes. Therefore, the protection of seagrass beds needs to be strengthened to promote the restoration of coastal fishery resources.
Key words: seagrass bed; fish; growth characteristics; seagrass resource
CHEN Qiming , LIU Songlin , ZHANG Chi , CUI Lijun , JIANG Zhijian , WU Yunchao , HUANG Xiaoping . Growth characteristics of four representative fishes and their responses to seagrass resource changes in typical tropical seagrass beds of Hainan Island[J]. Journal of Tropical Oceanography, 2020 , 39(5) : 62 -70 . DOI: 10.11978/2020007
表1 海南岛黎安港、新村湾和潭门港褐篮子鱼、点斑篮子鱼、四带牙鯻和细鳞鯻的样本量(单位: 尾)Tab. 1 Sample size for S. fuscessens, S. guttatus, P. quadrilineatus, and T. jarbua in the seagrass meadows of Li’an Port, Xincun Bay and Tanmen Port |
| 鱼种 | 黎安港 | 新村湾 | 潭门港 |
|---|---|---|---|
| 褐篮子鱼 | 33 | 33 | 75 |
| 点斑篮子鱼 | 31 | 30 | 40 |
| 四带牙鯻 | 40 | 77 | 26 |
| 细鳞鯻 | 47 | 25 | 31 |
表2 海南岛潭门港、新村湾和黎安港3个海草床海菖蒲的主要生物学指标(x ±均方误差, n=6~10)Tab. 2 Biological traits of Ehanlus acoroides in the three seagrass meadows of Tanmen Port, Xincun Bay and Li’an Port (x ± MSE, n = 6~10) |
| 海菖蒲海草生物学指标 | 潭门港 | 新村湾 | 黎安港 |
|---|---|---|---|
| 覆盖率/% | 24.60±2.75a | 47.60±3.04b | 61.20±6.97c |
| 茎枝密度/(株·m-2) | 147.20±5.99a | 214.40±13.00b | 256.00±25.30b |
| 海草根生物量/(g·m-2) | 192.26±81.12a | 222.59±28.74a | 539.19±227.24a |
| 海草茎生物量/(g·m-2) | 300.88±71.70a | 1300.85±182.35a | 2713.96±628.86b |
| 海草叶片生物量/(g·m-2) | 247.43±88.49a | 513.73±108.44a | 1121.80±127.07b |
注: 不同的小写字母(a、b、c)表示不同站位间的差异显著(S-N-K 检验, p < 0.05) |
图1 海南岛潭门港、新村湾和黎安港海草床褐篮子鱼(a)、点斑篮子鱼(b)、四带牙鯻(c)和细鳞鯻(d)体长和体质量之间的关系Fig. 1 Relationships of body mass and body length of S. fuscessens (a), S. guttatus (b), P. quadrilineatus (c), and T. jarbua (d) in the seagrass meadows of Li’an Port, Xincun Bay and Tanmen Port |
表3 海南岛潭门港、新村湾和黎安港海草床褐篮子鱼、点斑篮子鱼、四带牙鯻和细鳞鯻体长-体质量关系参数Tab. 3 Parameters of body length-mass relationship for S. fuscessens, S. guttatus, P. quadrilineatus and T. jarbua in the seagrass meadows of Li’an Port, Xincun Bay and Tanmen Port |
| 鱼种 | 采样地点 | 样本量 | 长度范围/cm | a | b | r2 | t |
|---|---|---|---|---|---|---|---|
| 褐篮子鱼 | 黎安港 | 33 | 6.7~23.0 | 0.01 | 3.23 | 0.99 | 2.94** |
| 新村湾 | 33 | 4.3~21.9 | 0.02 | 3.03 | 0.99 | 1.73ns | |
| 潭门港 | 75 | 6.4~27.8 | 0.04 | 2.76 | 0.98 | 5.07** | |
| 点斑篮子鱼 | 黎安港 | 31 | 7.1~21.0 | 0.03 | 3.05 | 0.97 | 3.78** |
| 新村湾 | 30 | 8.6~25.0 | 0.04 | 3.08 | 0.99 | 10.31** | |
| 潭门港 | 40 | 6.9~24.5 | 0.06 | 2.81 | 0.96 | 5.86** | |
| 四带牙鯻 | 黎安港 | 40 | 5.4~13.1 | 0.01 | 3.32 | 0.99 | 8.59** |
| 新村湾 | 77 | 4.4~12.6 | 0.01 | 3.20 | 0.99 | 10.33** | |
| 潭门港 | 26 | 8.4~9.8 | 0.01 | 2.95 | 0.95 | 6.52** | |
| 细鳞鯻 | 黎安港 | 47 | 4.9~14.0 | 0.02 | 3.17 | 0.97 | 8.57** |
| 新村湾 | 25 | 6.5~15.4 | 0.02 | 3.05 | 0.98 | 4.81** | |
| 潭门港 | 31 | 6.5~16.0 | 0.03 | 2.91 | 0.97 | 4.61** |
注: a, b为体长-体质量关系参数; r2为相关系数; t为b与3的差异性t检验值。ns表示b与3的差异不显著(p>0.05); *表示b与3的差异显著水平为p<0.05; **表示b与3的差异显著水平为p<0.01 |
表4 海南岛潭门港、新村湾和黎安港海草床间褐篮子鱼、点斑篮子鱼、四带牙鯻和细鳞鯻异速生长因子的差异性检验Tab. 4 Comparison of allometric factors of S. fuscessens, S. guttatus, P. quadrilineatus, and T. jarbua among the seagrass meadows of Li’an Port, Xincun Bay and Tanmen Port |
| 对比区域 | 褐篮子鱼 | 点斑篮子鱼 | 四带牙鯻 | 细鳞鯻 |
|---|---|---|---|---|
| 黎安港vs新村湾 | 0.95ns | 1.09ns | 0.13ns | 0.22ns |
| 黎安港vs潭门港 | 4.28* | 4.00* | 4.19* | 9.14** |
| 新村湾vs潭门港 | 3.37ns | 8.88** | 0.71ns | 3.25ns |
注: ns表示不同区域海草床间无显著差异(p>0.05); *表示不同区域海草床间差异显著水平为p<0.05; **表示不同区域海草床间差异显著水平为p<0.01 |
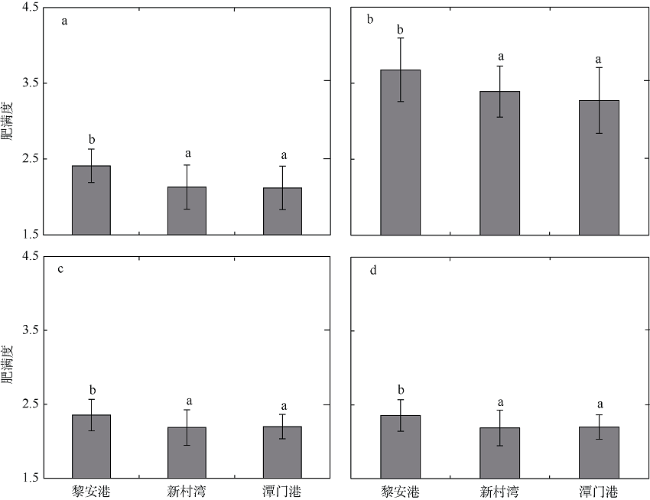
图2 海南岛黎安港、新村湾和潭门港海草床褐篮子鱼(a)、点斑篮子鱼(b)、四带牙鯻(c)和细鳞鯻(d)肥满度的差异数据柱上不同小写字母(a, b)表示不同区域海草床存在显著性差异(S-N-K检验, p<0.05) Fig. 2 Variation in relative fatness of S. fuscessens (a), S. guttatus (b), P. quadrilineatus (c), and T. jarbua (d) in the seagrass meadows of Li’an Port, Xincun Bay and Tanmen Port. Different capital letters over the bars indicate significant difference among different seagrass meadows (S-N-K test, p < 0.05) |
表5 全球不同海域褐篮子鱼、点斑篮子鱼、四带牙鯻和细鳞鯻异速生长因子的差异Tab. 5 Comparison of allometric factors of S. fuscessens, S. guttatus, P. quadrilineatus, and T. jarbua in global coastal areas |
| 研究区域 | 褐篮子鱼 | 点斑篮子鱼 | 四带牙鯻 | 细鳞鯻 | 参考文献 |
|---|---|---|---|---|---|
| 菲律宾, 巴拉望岛 | 2.51 | Palla等(2018) | |||
| 菲律宾, 普哈达湾 | 3.01 | Jumawan-Nanual等(2008) | |||
| 新喀里多尼亚 | 3.01 | Letourneur等(1998) | |||
| 菲律宾, 达沃湾 | 3.01 | 2.89 | Gumanao等(2016) | ||
| 菲律宾, 本田湾 | 2.95 | Jumawan-Nanual等(2008) | |||
| 日本, 冲绳海域 | 3.00 | Database of IGFA (2009) | |||
| 土耳其, 地中海 | 2.96 | Taskavak (2001) | |||
| 泰国, 罗勇府 | 2.88 | Yamagawa (1994) | |||
| 南非, 河口 | 2.94 | Harrison (2001) | |||
| 也门, 亚丁湾 | 2.99 | Lavergne等(2013) | |||
| 中国北部湾 | 2.76 | Wang等(2011) | |||
| 中国湛江沿海 | 2.88 | 张健东等(2002) | |||
| 中国香港西、南及东部水域 | 3.11 | Perkins等(2019) | |||
| 中国海南岛海域 | 3.02 | 3.02 | 3.14 | 3.04 | 本研究 |
| [1] |
樊敏玲, 黄小平, 张大文, 等, 2011. 海南新村湾海草床主要鱼类及大型无脊椎动物的食源[J]. 生态学报, 31(1):31-38.
|
| [2] |
黄晓璇, 2010. 青岛近海方氏云鳚(Enedrias fangi Wang et Wang)渔业生物学初步研究[D]. 青岛: 中国海洋大学: 9-10.
|
| [3] |
李忠炉, 2011. 黄渤海小黄鱼、大头鳕和黄鮟鱇种群生物学特征的年际变化[D]. 青岛: 中国科学院研究生院(海洋研究所):11-19.
|
| [4] |
刘松林, 江志坚, 吴云超, 等, 2015. 海草床育幼功能及其机理[J]. 生态学报, 35(24):7931-7940.
|
| [5] |
罗秉征, 1966. 浙江近海大黄鱼的季节生长[J]. 海洋与湖沼, 8(2):121-139.
|
| [6] |
张健东, 宋蓓玲, 陈刚, 2002. 细鳞鯻年龄与生长的研究[J]. 海洋科学, 26(7):63-67.
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
IGFA, 2009. Database of IGFA angling records until 2009[DB]. IGFA, Fort Lauderdale, USA. http://wrec.igfa.org/.
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}