
Journal of Tropical Oceanography >
Effects of salinity changes on serum and kidney immune status associated with injection of Aeromonas hydrophila in Scatophagus argus
Copy editor: LIN Qiang
Received date: 2020-03-09
Request revised date: 2020-04-07
Online published: 2020-04-09
Supported by
National Natural Science Foundation of China(41806177)
National Natural Science Foundation of China(41976108)
China Postdoctoral Science Foundation(2019M653010)
Copyright
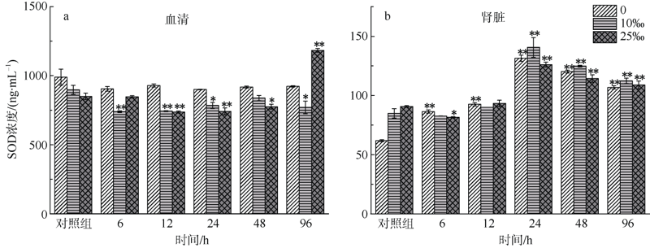
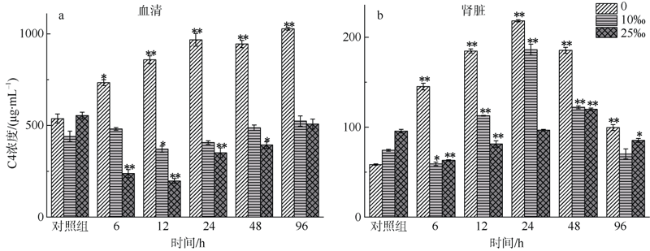
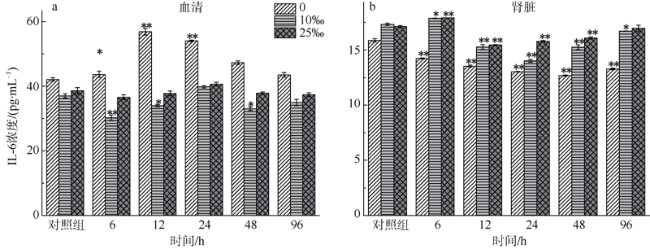
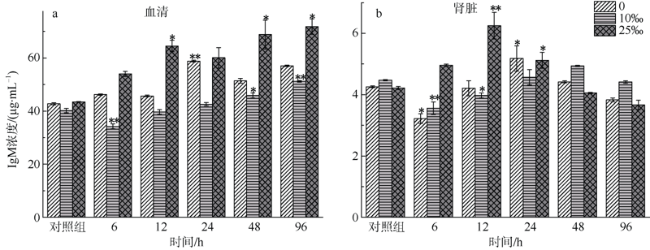
The salinity in coastal waters fluctuates frequently due to seasonal rainfall and ocean currents. Salinity change makes fish more susceptible to pathogens, leading to disease and death. As the main lymphoid tissue of teleosts, kidney is closely related to fish’s immune function. This study aims to explore effects of environmental salinity on the kidney immune function of Scatophagus argus. In order to analyze the relationship between the change of environmental salinity and immune situation in fish’s kidney after Aeromonas hydrophila injection, we compared the changes of superoxide dismutase (SOD) concentration in serum and kidney, and immune-related parameters Complement 4 (C4), Interleukin-6 (IL-6), and Immunoglobulin M (IgM) concentrations in kidney before and after infection within 96 h of each salinity group. Our results show that except for 96 hpi, SOD concentrations in the serum and kidney tissues of the freshwater group and low-salinity (?‰) group were higher than those of the 25‰ salinity group (P<0.01), and that the maximum difference between low-salinity group and 25‰ salinity group was about 150 ng·mL-1. The concentration of C4 in the serum and kidney of the freshwater group and low-salinity group was about 100-600 μg·mL-1 higher than that of the 25‰ salinity group within 96 hpi (P<0.01). The concentration of IL-6 in the kidney tissues of S. argus in the 25‰ salinity group was higher than that in the freshwater group and low-salinity group (P<0.01), although IL-6 concentrations in kidney of the three groups were significantly lower than that of the control group (14.2±0.1 pg·mL-1, in freshwater; 17.9±0.0 pg·mL-1, in low-salinity; 17.9±0.0 pg·mL-1, in 25‰ salinity) at 96 hpi (P<0.01), while the serum IL-6 concentration in the freshwater group was significantly increased to 56.9±1.0 pg·mL-1 (P<0.01). The serum and kidney IgM levels of fish in the 25‰ salinity grop (71.8±2.9 μg·mL-1 and 6.3±0.4 μg·mL-1) were about 1-20 g·mL-1 higher than those of the freshwater group and low-salinity group (P<0.05). The time of IgM production in serum and kidney tissues of the 25‰ salinity group (12 hpi) was earlier than that of the freshwater group and low-salinity group (24 hpi). In summary, the 25‰ salinity group suffered less damage after bacterial infection, and the immune status of kidney and serum was better than that of the freshwater group and low-salinity group; and the immune response was faster in the 25‰ salinity group. Therefore, we speculate that a decrease of salinity would lead to a decrease of kidney immune state of S. argus. This research provides a reference value for S. argus mariculture.
Key words: Scatophagus argus; salinity; Aeromonas hydrophila; immunity; kidney
LU Mengying , SU Maoliang , ZHANG Junbin . Effects of salinity changes on serum and kidney immune status associated with injection of Aeromonas hydrophila in Scatophagus argus[J]. Journal of Tropical Oceanography, 2021 , 40(3) : 114 -123 . DOI: 10.11978/2020026
图1 不同盐度金钱鱼注射嗜水气单胞菌后96h内血清(a)和肾脏(b)组织中SOD浓度(n=5)**P < 0.01; * P < 0.05(与对照组相比较) Fig.1 Concentrations of SOD in serum (a) and kidney (b) among three salinity experimental groups after A. hydrophila injection at 6, 12, 24, 48, and 96 h (n=5). **P < 0.01; *P < 0.05 (compared to the control group) |
图2 不同盐度金钱鱼注射嗜水气单胞菌后96h内血清(a)和肾脏(b)中C4浓度变化(n=5)**P < 0.01; * P < 0.05(与对照组相比较) Fig. 2 Concentrations of C4 in serum (a) and kidney (b) among three salinity experimental groups after A. hydrophila injection at 6, 12, 24, 48, and 96 h (n=5). **P < 0.01; *0.01 < P < 0.05 (compared to the control group) |
图3 不同盐度金钱鱼注射嗜水气单胞菌后96h内血清和肾脏中IL-6浓度变化(n=5)**P < 0.01; *P < 0.05(与对照组相比较) Fig. 3 Concentrations of IL-6 in serum (a) and kidney (b) among three salinity experimental groups after A. hydrophila injection at 6, 12, 24, 48, and 96 h (n=5). **P < 0.01; * P < 0.05 (compared to the control group) |
图4 不同盐度金钱鱼注射嗜水气单胞菌后96h内血清(a)和肾脏(b)中IgM浓度变化(n=5)**P < 0.01; *P < 0.05(与对照组相比较) Fig. 4 Concentrations of IgM in serum (a) and kidney (b) among three salinity experimental groups after A. hydrophila injection at 6, 12, 24, 48, and 96 h (n=5). **P < 0.01; * P < 0.05 (compared to the control group) |
| [1] |
何杰, 强俊, 徐跑, 等, 2014. 低温驯化下4种不同品系罗非鱼血清皮质醇与免疫相关指标的变化[J]. 中国水产科学, 21(2):266-274.
|
| [2] |
侯月娥, 冯娟, 杨清华, 等, 2010. 鱼类免疫球蛋白的研究进展[J]. 黑龙江畜牧兽医, 0(9):37-39.
|
| [3] |
梁雪梅, 2018. 催乳素在金钱鱼盐度适应过程中的机理研究[D]. 上海: 上海海洋大学.
|
| [4] |
杨尉, 陈华谱, 江东能, 等, 2018. 金钱鱼生物学及繁养殖技术研究进展[J]. 生物学杂志, 35(5):104-108.
|
| [5] |
张庆华, 马文元, 陈彪, 等, 2016. 嗜水气单胞菌引致的金钱鱼细菌性疾病[J]. 水产学报, 40(4):634-643.
|
| [6] |
张媛媛, 宋理平, 2018. 鱼类免疫系统的研究进展[J]. 河北渔业, (2):49-56.
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}