
Journal of Tropical Oceanography >
Phytoplankton responses to Dan’ao River estuary water enrichment in terms of size structure and community composition*
Copy editor: YIN Bo
Received date: 2020-04-14
Request revised date: 2020-06-01
Online published: 2020-06-01
Supported by
Natural Science Foundation of China(31971432)
Natural Science Foundation of China(41506161)
Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0405)
Guangdong Marine Economy Promotion Projects Fund(GDOE[2019]A32)
Science and Technology Planning Project of Guangdong Province, China(2017B0303014052)
Innovation Academy of South China Sea Ecology and Environmental Engineering, Chinese Academy of Sciences(ISEE2018PY01)
Copyright
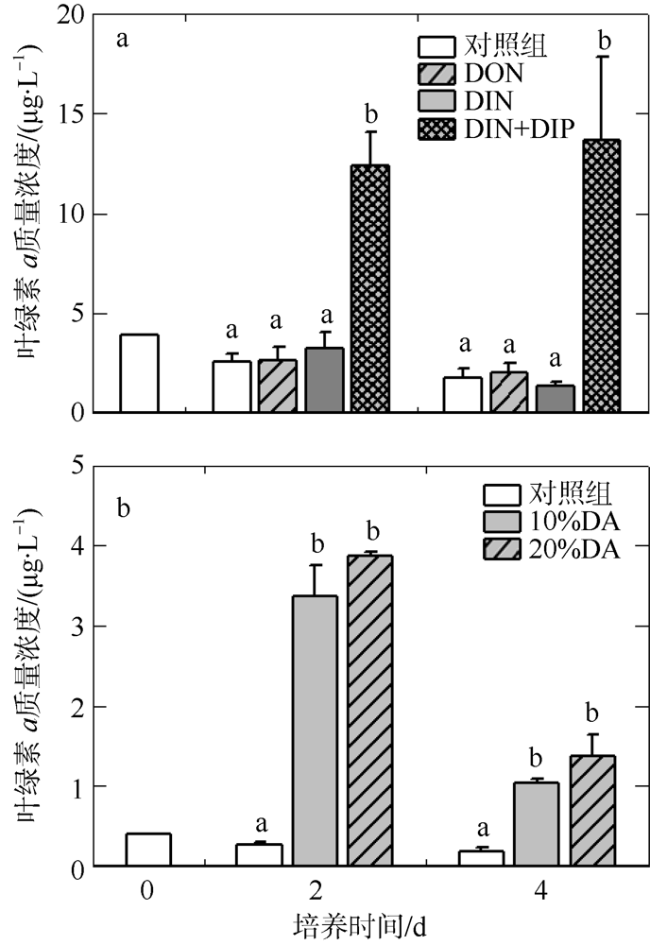
Daya Bay is experiencing ecological problems such as eutrophication and frequent algal bloom under the pressure of intense anthropogenic activities. The Dan’ao River is the largest rive entering Daya Bay. We found the concentrations of total dissolved nitrogen and phosphorus at the Dan’ao River estuary reached 85.3 and 1.5 μmol·L-1, respectively. In situ enrichment experiments were conducted to explore the effect of estuary water from the Dan’ao River on phytoplankton community. Our results showed filtered water (0.2 μm) from the Dan’ao River estuary promoted total Chlorophyll a (Chl a) and phytoplankton abundance, and shifted the dominated Chl a size structure to smaller size (0.7 ~ 20 μm), and also increased the relative abundance of dinoflagellates (Scrippsiella trochoidea and Prorocentrum spp.). The phytoplankton size structure and species composition changed in the same pattern under the addition of dissolved organic nitrogen (DON) of urea. Although combined addition of dissolved inorganic nitrogen and phosphorus increased total Chl a and abundance, the phytoplankton size structure and dinoflagellate abundance did not show significant changes. Our results demonstrate that DON may be the causative nutrient component that leads to miniaturization of the phytoplankton community and promotes the growth of dinoflagellate by comparing the enrichment results of estuary water to N and P nutrient enrichments. These results indicate the significance of DON for eutrophication and harmful algal blooms in Daya Bay, which should be included in management strategies of coastal water eutrophication.
Key words: Daya Bay; Dan’ao River; urea; phytoplankton community; size
XIANG Chenhui , LIU Jiaxing , KE Zhixin , ZHOU Linbin , TAN Yehui . Phytoplankton responses to Dan’ao River estuary water enrichment in terms of size structure and community composition*[J]. Journal of Tropical Oceanography, 2021 , 40(2) : 49 -60 . DOI: 10.11978/2020040
表1 现场加富培养试验设计Tab. 1 Design of nutrient additions of in situ enrichment experiments |
| 试验名称 | 组名 | 加富内容 |
|---|---|---|
| 试验1: 氮、磷营养加富试验 | 对照组 | 无 |
| DON组 | 氮浓度为14μmol·L-1的尿素[CO(NH2)2]溶液 | |
| DIN组 | 氮浓度为14μmol·L-1的NaNO3溶液 | |
| DIN+DIP组 | 14μmol·L-1 NaNO3溶液+ 1μmol·L-1 NaH2PO4溶液 | |
| 试验2: 河口水加富试验 | 对照组 | 无DA |
| 10% DA组 | 0.2L DA | |
| 20% DA组 | 0.4L DA |
注: DON: 溶解性有机氮; DIN: 溶解性无机氮; DIP: 溶解性无机磷; DA: 经0.2µm聚碳酸酯滤膜(Millipore, 47mm)过滤后的淡澳河口S3站位表层水。每个培养体系为2L |
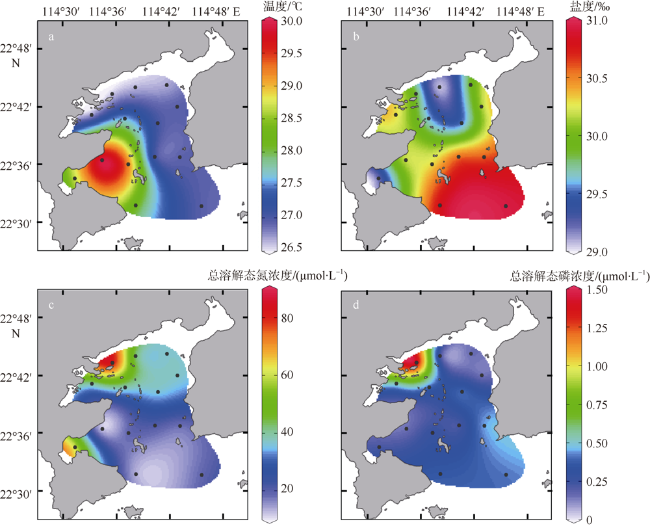
图2 2016年10月大亚湾表层海水温度(a)、盐度(b)、总溶解态氮浓度(c)和总溶解态磷浓度(d)水平分布审图号: GS(2019)4342 Fig. 2 Surface water temperature (a), salinity (b), total dissolved nitrogen (c), and total dissolved phosphorus (d) in Daya Bay in October 2016 |
表2 2016年秋季大亚湾中心(S10)和淡澳河口(S3)海水理化和生物特征Tab. 2 Initial physicochemical conditions and phytoplankton community composition at central bay (S10) and Dan’ao River estuary (S3) for enrichment experiments in Daya Bay in October 2016 |
| 站位 | 温度/℃ | 盐度/‰ | TDN浓度/ (μmol·L-1) | DIN : DON | TDP浓度/ (μmol·L-1) | DIP : DOP | 叶绿素a质量浓度/ (µg·L-1) | 浮游植物丰度/ (个·L-1) |
|---|---|---|---|---|---|---|---|---|
| S10 | 29.3 | 30.2 | 19.5 | 90 : 10 | 0.3 | 31 : 69 | 3.9 | 3.4×105 |
| S3 | 26.6 | 30.0 | 85.3 | 59 : 41 | 1.5 | 63 : 37 | 3.5 | 5.8×104 |
注: TDN: 总溶解态氮; DIN: 溶解性无机氮; DON: 溶解性有机氮; TDP: 总溶解态磷; DIP: 溶解性无机磷; DOP: 溶解性有机磷 |
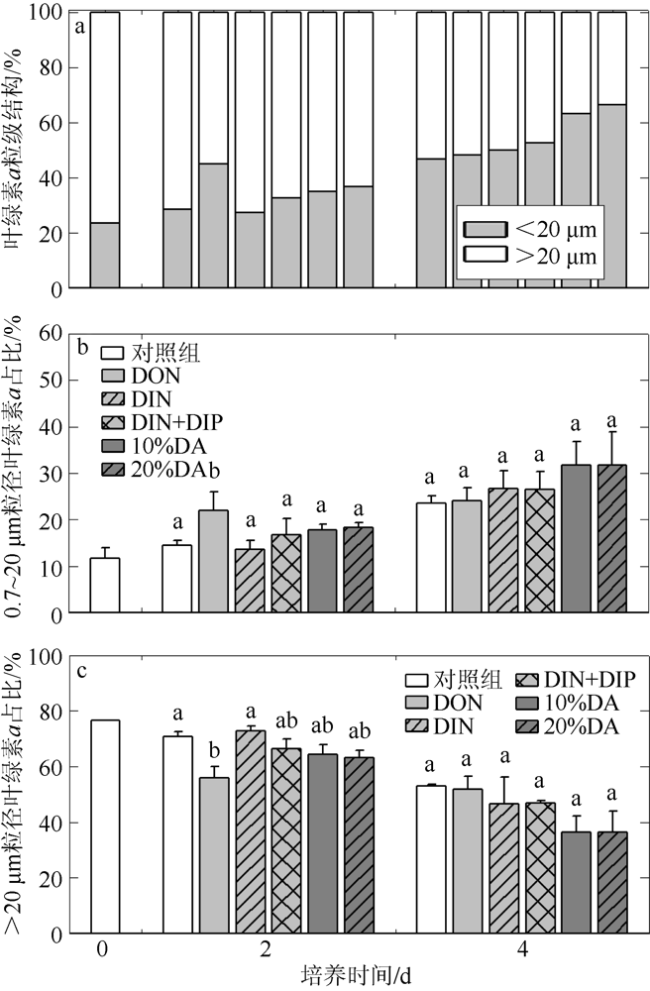
图4 叶绿素a粒级结构(a)和分级叶绿素a贡献量(b, c)对氮、磷营养, 淡澳河河口水加富的响应DON: 溶解性有机氮; DIN: 溶解性无机氮; DIP: 溶解性无机磷; DA: 经0.2µm聚碳酸酯滤膜(Millipore, 47mm)过滤后的淡澳河口S3站位表层水。图中不同字母表示该指标在同一天内的两个试验组之间具有显著差异(p<0.05); 图a组别与图b组别相对应 Fig. 4 Response of chlorophyll a (Chl a) size structure (a) and proportion of size-fractioned Chl a (b, c) concentration to nitrogen, phosphorus, and estuary water enrichment of Dan’ao River |
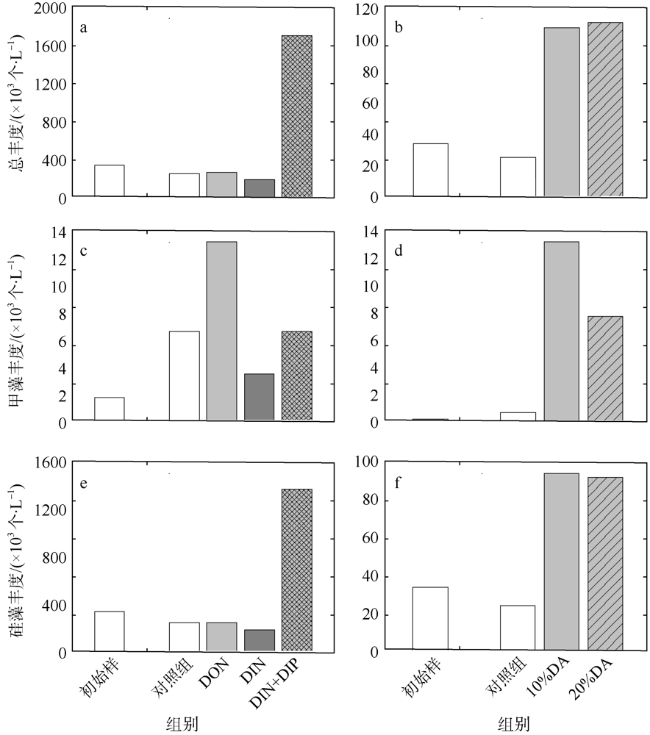
图5 培养至第2天时浮游植物总丰度(a, b)、甲藻(c, d)和硅藻(e, f)丰度对氮、磷营养(a, c, e)和淡澳河河口水(b, d, f)加富的响应DON: 溶解性有机氮; DIN: 溶解性无机氮; DIP: 溶解性无机磷; DA: 经0.2µm聚碳酸酯滤膜(Millipore, 47mm)过滤后的淡澳河口S3站位表层水 Fig. 5 Responses of phytoplankton total abundance (a, b), dinoflagellate abundance (c, d) and diatom abundance (e, f) to nitrogen and phosphorus (a, c, e), and to estuary water enrichment of Dan’ao River (b, d, f) |
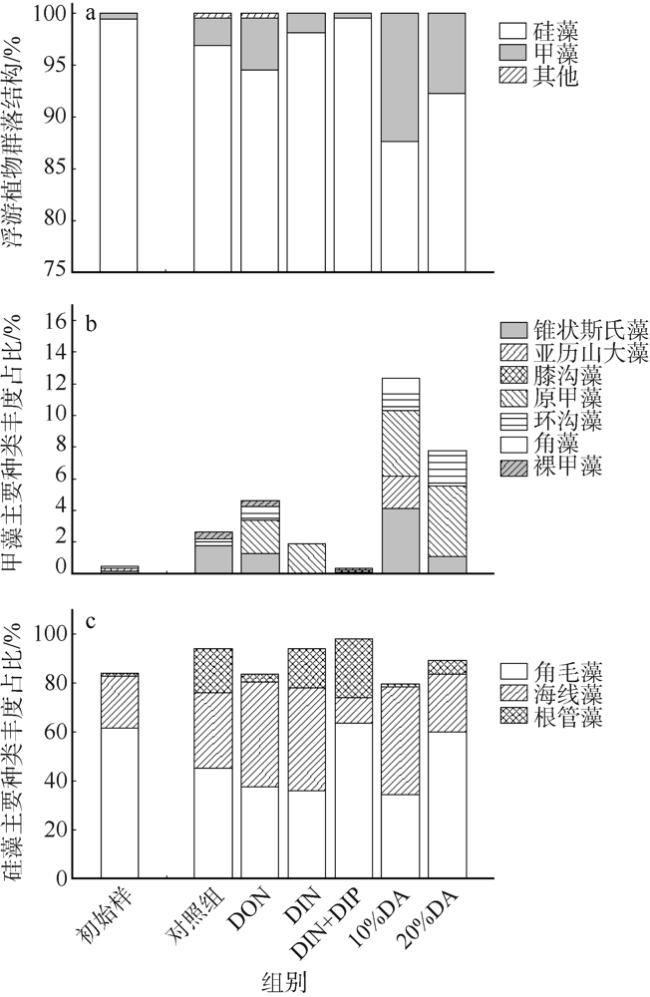
图6 培养至第4天时浮游植物群落结构(a), 甲藻(b)和硅藻(c)主要类群对氮、磷营养和淡澳河河口水加富的响应DON: 溶解性有机氮; DIN: 溶解性无机氮; DIP: 溶解性无机磷; DA: 经0.2µm聚碳酸酯滤膜(Millipore, 47mm)过滤后的淡澳河口S3站位表层水 Fig. 6 Responses of phytoplankton community composition (a), dinoflagellate dominant species (b) and diatom dominant species (c) to nitrogen and phosphorus and to estuary water enrichment of Dan’ao River |
表3 各试验组营养盐初始浓度Tab. 3 Initial nutrient concentrations in each treatment |
| 组别 | DIN浓度/(μmol·L-1) | DON浓度/(μmol·L-1) | DIP浓度/(μmol·L-1) | DOP浓度/(μmol·L-1) | SiO32-浓度/(μmol·L-1) |
|---|---|---|---|---|---|
| 对照 | 17.6 | 1.9 | 0.1 | 0.2 | 2.5 |
| DON | 17.6 | 15.9 | 0.1 | 0.2 | 2.5 |
| DIN | 31.6 | 1.9 | 0.1 | 0.2 | 2.5 |
| DIN+DIP | 31.6 | 1.9 | 1.1 | 0.2 | 2.5 |
| 10% DA | 20.8 | 5.3 | 0.2 | 0.2 | 4.4 |
| 20% DA | 24.1 | 8.6 | 0.3 | 0.3 | 6.4 |
注: DIN: 溶解性无机氮; DON: 溶解性有机氮; DIP: 溶解性无机磷; DOP: 溶解性有机磷; DA, 经0.2µm聚碳酸酯滤膜(Millipore, 47mm)过滤后的淡澳河口S3站位表层水 |
| [1] |
胡章喜, 徐宁, 段舜山, 等, 2010. 尿素对中国近海3种典型赤潮藻生长的影响[J]. 环境科学学报, 30(6):1265-1270.
|
| [2] |
桓清柳, 庞仁松, 周秋伶, 等, 2016. 深圳近岸海域氮、磷营养盐变化趋势及其与赤潮发生的关系[J]. 海洋环境科学, 35(6):908-914.
|
| [3] |
黄小平, 黄良民, 宋金明, 等, 2019. 营养物质对海湾生态环境影响的过程与机理[M]. 北京: 科学出版社: 16-17(in Chinese).
|
| [4] |
赖海燕, 徐宁, 段舜山, 2011. 大亚湾裸甲藻种群动态及其关键调控因子[J]. 生态环境学报, 20(3):505-510.
|
| [5] |
李佳俊, 沈萍萍, 谭烨辉, 等, 2015. 南海东北部浮游植物对氮、磷加富的响应及与不同水团的关系[J]. 海洋学报, 37(10):88-99.
|
| [6] |
李涛, 2007. 大亚湾水域浮游植物对环境变化的响应研究[D]. 广州: 中国科学院南海海洋研究所.
|
| [7] |
刘冬燕, 赵建夫, 马利民, 等, 2004. 粒径分级叶绿素a对富营养水体生物修复的响应[J]. 生态学报, 24(11):2477-2483.
|
| [8] |
任秀文, 姜国强, 刘爱萍, 等, 2013. 大亚湾主要入海河流污染物通量估算研究[C]// 2013中国环境科学学会学术年会. 昆明: 2912-2921 (in Chinese).
|
| [9] |
宋淑华, 2009. 大亚湾浮游植物群落结构与赤潮藻分子鉴定研究[D]. 广州: 暨南大学.
|
| [10] |
王朝晖, 陈菊芳, 徐宁, 等, 2001. 大亚湾裸甲藻种群的季节变化与环境条件的关系[J]. 生态学报, 21(11):1825-1832.
|
| [11] |
王友绍, 王肇鼎, 黄良民, 2004. 近20年来大亚湾生态环境的变化及其发展趋势[J]. 热带海洋学报, 23(5):85-95.
|
| [12] |
肖咏之, 齐雨藻, 王朝晖, 等, 2001. 大亚湾海域锥状斯氏藻赤潮及其与孢囊的关系[J]. 海洋科学, 25(9):50-54.
|
| [13] |
谢福武, 刘华雪, 黄洪辉, 等, 2018. 大亚湾浮游植物粒级结构对温排水和营养盐输入的响应[J]. 热带海洋学报, 37(3):55-64.
|
| [14] |
徐宁, 陈菊芳, 王朝晖, 等, 2001. 广东大亚湾藻类水华的动力学分析Ⅱ. 藻类水华与营养元素的关系研究[J]. 环境科学学报, 21(4):400-404.
|
| [15] |
易斌, 陈凯彪, 周俊杰, 等, 2018. 2009年至2016年华南近海赤潮分布特征[J]. 海洋湖沼通报, (2):23-31.
|
| [16] |
张淑雯, 张云, 欧林坚, 等, 2012. 6种赤潮甲藻对荧光标记藻类的吞噬行为研究[J]. 海洋与湖沼, 43(3):602-608.
|
| [17] |
张云, 黄凯旋, 欧林坚, 等, 2014. 大亚湾海域尿素与浮游生物脲酶活性研究[J]. 热带海洋学报, 33(1):90-96.
|
| [18] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. GB/T 12763.4-2007 海洋调查规范第4部分: 海水化学要素调查[S]. 北京: 中国标准出版社.
GENERAL ADMINISTRATION OF QUALITY SUPERVISION, INSPECTION AND QUARANTINE OF THE PEOPLE'S REPUBLIC OF CHINA, STANDARDIZATION ADMINISTRATION, 2008. GB/T 12763.4-2007 Specifications for oceanographic survey-Part 4: Survey of chemical parameters in sea water[S]. Beijing: Standards Press of China (in Chinese).
|
| [19] |
朱艾嘉, 黄良民, 许战洲, 2008. 氮、磷对大亚湾大鹏澳海区浮游植物群落的影响Ⅰ. 叶绿素a与初级生产力[J]. 热带海洋学报, 27(1):38-45.
|
| [20] |
朱艾嘉, 黄良民, 林秋艳, 等, 2009. 氮、磷对大亚湾大鹏澳海区浮游植物群落的影响: Ⅱ 种类组成[J]. 热带海洋学报, 28(6):103-111.
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}