
Journal of Tropical Oceanography >
Morphological and molecular study on marine green alga, Halimeda velasquezii, first recorded in Hainan Island
Copy editor: LIN Qiang
Received date: 2020-02-24
Request revised date: 2020-05-14
Online published: 2020-06-22
Supported by
Financial Fund of the Ministry of Agriculture and Rural Affairs of China(NFZX2018)
National Non-profit Institute Research Grant of CATAS-ITBB from Chinese Government(1630052019010)
National Non-profit Institute Research Grant of CATAS-ITBB from Chinese Government(1630052020016)
Copyright
Samples of Halimeda were collected from the intertidal zone of Fengjia Bay in Hainan Island during the field survey in April 2019. Detailed morphological structure observation and molecular systematics analysis were conducted. The results showed that the sample was identified as Halimeda velasquezii W.R. Taylor. We found the following combination of the characters: the thalli is green, calcified, erect from a single small stupose holdfast; segments are transversely oval to reniform linking together; utricles form in surface view is round or a little compressed; filaments inside are long tubular; medullary filaments fuse for a short distance in pairs in common; subcortical filaments loosely branch, and are not utriculiform. The rbcL gene analysis showed that the sample was clustered on the same branch as that from the Indian Ocean-Pacific Ocean region, whose bootstrap value and sequence divergence were 100 and 9 bp (0.69%), respectively. The species was the first recorded in Hainan Island. The results should enrich our knowledge of Chinese species’ morphology and extend their geographical distribution. They should be of great significance to understand the biodiversity and distribution characteristics of algal resources in the waters of Hainan Island.
Key words: Halimeda velasquezii; Hainan Island; classification; morphology; rbcL gene
MENG Tian , CHEN Zuo , ZHU Jun , ZOU Xiaoxiao , FU Qingyan , BAO Shixiang . Morphological and molecular study on marine green alga, Halimeda velasquezii, first recorded in Hainan Island[J]. Journal of Tropical Oceanography, 2020 , 39(6) : 114 -121 . DOI: 10.11978/2020021
表1 分子序列分析所用海藻种类、采集地点及基因登录号列表Tab. 1 List of species used in this study, collection locations, and GenBank accession numbers |
| 物种 | 采集地点 | rbcL序列登录号 |
|---|---|---|
| Halimeda velasquezii Taylor | 印度洋-太平洋地区 | FJ624507.1 |
| H. borneensis W.R.Taylor | 澳大利亚新厄里多尼亚岛 | AB619273.1 |
| 澳大利亚新厄里多尼亚岛 | AB700353.1 | |
| H. cylindracea Decaisne | 印度洋-太平洋地区 | FJ624510.1 |
| H. gracilis Harvey ex J.Agardh | 大西洋地区 | FJ624493.1 |
| H. heteromorpha N'Yeurt | 印度洋-太平洋地区 | FJ624511.1 |
| H. kanaloana Vroom | 印度洋-太平洋地区 | FJ624512.1 |
| H. macroloba Decaisne | 印度洋-太平洋地区 | FJ624513.1 |
| H. melanesica Valet | 印度洋-太平洋地区 | FJ624515.1 |
| Udotea flabellum (J.Ellis & Solander) M.Howe | 加勒比海地区 | MG784872.1 |
| U. argentea Zanardini | 澳大利亚 | NC_039527.1:45455-46882 |
| Boodleopsis pusilla (Collins) W.R.Taylor, A.B.Joly & Bernatowicz | 澳大利亚 | NC_039520.1:24118-25545 |
| Chlorodesmis sp. Tenija_051113A | 日本冲绳 | AB619306.1 |
| 日本冲绳 | AB619308.1 | |
| Chlorodesmis sp. Tenija_050807A | 日本冲绳 | AB619311.1 |
| Caulerpa racemosa (Forsskål) J.Agardh | 日本Tohwa-cho | AB038486.1 |
| C. subserrata Okamura | 日本神户 | AB054015.1 |
| C. taxifolia (M.Vahl) C.Agardh | 日本神户 | AB054016.1 |
图1 未氏仙掌藻孢子体的外观形态与解剖结构观察a. 外观形态; b—d. 光学显微镜下结构观察, C示皮层, ms示髓丝, pu示初级囊胞; e—f. 体式显微镜下结构观察, nf示节点处融合, g示配子囊 Fig. 1 Morphological appearance and anatomical structure of H. velasquezii. a) morphological appearance; (b-d) anatomical structure by light microscope, C: cortex; ms: medullary filaments; pu: primary utriculiform; (e-f) anatomical structure by stereomicroscope, nf: node fusion; g: gametangium |
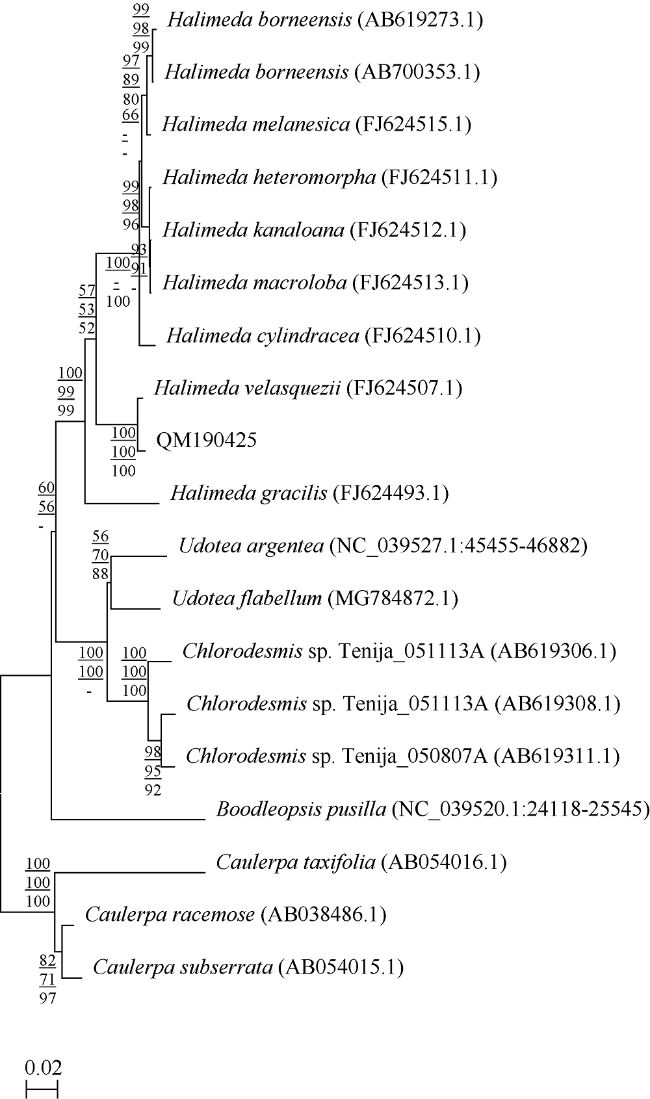
图2 基于rbcL序列所构建的NJ系统发育树节点的数值表示自展支持值, 从上至下分别表示NJ、ML、MP, 自展值只显示大于50 Fig. 2 Neighbor joining (NJ) tree built based on rbcL sequences. The numbers on the branch points represent bootstrap values (the one larger than 50 is shown), which from top to bottom represents NJ, ML, and MP respectively |
表2 本研究样本与已报道的属内形态相似种的特征比较(Dijoux et al, 2012)Tab. 2 Morphological comparison of the sample and similar species recorded before (Dijoux et al, 2012) |
| 样本与物种 | 形态特征 | |||||
|---|---|---|---|---|---|---|
| 藻体形状 | 节片形状 | 节片尺寸 | 固着器 | 节点融合 | 囊胞表面观 | |
| QM190425 | 直立 | 全缘, 横卵形或肾形 | 长(4~10mm) <宽(5~15mm) | 单一, 成簇细丝状 | 2~5根丝体局部 侧面融合 | 圆形或椭圆形 |
| H. velasquezii | 直立或下垂 | 边缘平整, 宽椭圆形或肾形 | 长(2~7mm) <宽(3~10mm) | 单一, 丝状假根束 | 2~3根丝体局部融合 | 圆形或多边形 |
| H. discoidea | 匍匐生长 | 边缘平整, 肾形或三叶形, 常有中肋 | 长(4~8mm) <宽(6~20mm) | 多个分散的假根固着器 | 2~5根丝体局部 侧面融合 | 多边形 |
| H. opuntia | 匍匐生长, 形成矮密藻丛 | 边缘平整, 肾形, 有时有中肋 | 长(3~7mm) <宽(4~8mm) | 多个分散的假根固着器 | 2根局部融合, 很少有3根融合 | 多边形, 有时圆形 |
| H. macroloba | 直立 | 边缘平整, 盘形或肾形 | 长(8~20mm) <宽(14~30mm) | 球根状 | 融合在一起, 中间有气孔 | 圆形或多边形 |
| H. gracilis | 匍匐生长, 分枝 稀疏 | 边缘平整, 三叶形、宽卵形或窄楔形 | 长(2~6mm)=宽(1~6mm) | 多个分散的假根固着器 | 2根(很少3根 ) 完全融合 | 多边形, 有时圆形 |
| H. heteromorpha | 直立, 丛生 | 边缘平整, 三叶形、 肾形 | 长(3~8mm) >宽(1~10mm) | 单一, 丝状假根束 | 融合在一起, 中间无气孔 | 多边形 |
| H. cylindracea | 直立 | 圆柱状或三叶状 | 长(1~9mm)=宽(1~8mm) | 球根状 | 融合在一起, 中间有气孔 | 圆形或多边形 |
| H. borneensis | 直立 | 边缘平整, 三叶形或盘形 | 长(4~11mm)=宽(4~14mm) | 球根状 | 融合在一起, 中间有气孔 | 圆形或多边形 |
| H. kanaloana | 直立 | 边缘平整, 宽三叶形或扇形 | 长(12mm) <宽(15~20mm) | 球根状 | 融合在一起, 中间有气孔 | 多边形 |
| H. melanesica | 直立 | 边缘平整, 三叶形或楔形 | 长(3~5mm)=宽(4~5mm) | 单一, 丝状假根束 | 融合在一起, 中间有小气孔 | 多边形 |
| [1] |
丁兰平, 黄冰心, 栾日孝, 2015. 中国海洋绿藻门新分类系统[J]. 广西科学, 22(2):201-210.
|
| [2] |
董美龄, 曾呈奎, 1980. 西沙群岛海产绿藻研究Ⅱ[J]. 海洋科学集刊, 17:1-7.
|
| [3] |
陆保仁, 曾呈奎, 董美龄 , 等, 1991. 南沙群岛海区褐藻和绿藻的研究Ⅰ[M] //中国科学院南沙综合科学考察队. 南沙群岛及其邻近海区海洋生物研究论文集. 北京: 海洋出版社: 1-14.
|
| [4] |
姚雪, 于丹, 王绪敏 , 等, 2011. 大型海洋藻类DNA条形码技术的开发与应用[J]. 今日科苑, ( 2):161-162.
|
| [5] |
曾呈奎, 董美龄, 1978. 西沙群岛海产绿藻的研究I[J]. 海洋科学集刊, 12:41-50.
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}