
Journal of Tropical Oceanography >
Functional study of coupling protein CheV and CZB domain of chemoreceptors in the Epsilon-proteobacteria chemotaxis signaling pathway
Copy editor: YIN Bo
Received date: 2020-05-26
Request revised date: 2020-06-29
Online published: 2020-07-29
Supported by
National Science Foundation of China(31870064)
Copyright
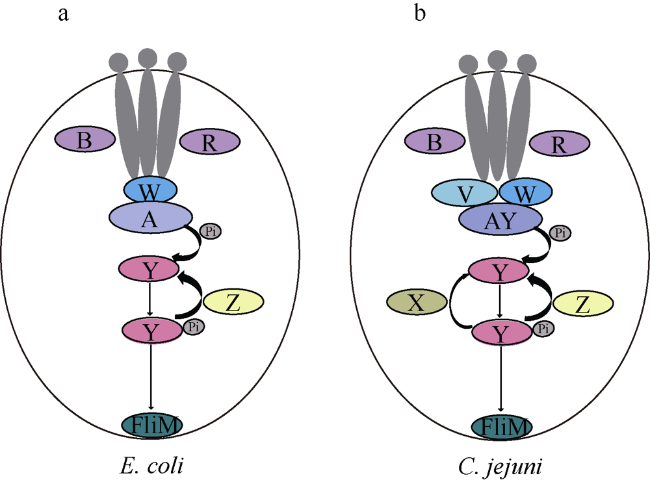
Epsilon-proteobacteria is widely distributed, from deep-sea hydrothermal vent to surface sea water, from free-living environment to host-associated one. Chemotaxis plays an important role in bacteria survival through sensing and responding to environmental changes. Thus, some bacteria have evolved into many complex and diverse chemotaxis systems. Epsilon-proteobacteria can adapt to different environments; especially, some species can survive in the deep-sea extreme environments including hydrothermal vent and cold seep, whose chemotaxis system may have special characteristics. Bioinformatics analyses by BlastP and MIST database revealed that most Epsilon-proteobacteria species in the deep-sea hydrothermal vent have F3 type chemotaxis system. They all contain a single copy of CheV, which is a double domain fusion protein. Besides, a unique domain, CZB (C-terminal Zinc-Binding) domain, exists in chemoreceptors of deep-sea Epsilon- proteobacteria. A CZB-like domain is also identified in Epsilon-proteobacteria. Using the model strain Campylobacter jejuni 81-176, we confirmed that CheV can interact with all chemoreceptors with MA domain by bacterial two-hybrid experiments. Additionally, we demonstrated that CZB-like domain cannot bind Zn by ICP-Mass, but it can promote the interaction between chemoreceptor Tlp9 and CheV.
Key words: Epsilon-proteobacteria; chemotaxis; CheV; CZB domain
LIU Yugeng , MAO Yingjin , ZHANG Canchuan , GAO Beile . Functional study of coupling protein CheV and CZB domain of chemoreceptors in the Epsilon-proteobacteria chemotaxis signaling pathway[J]. Journal of Tropical Oceanography, 2021 , 40(2) : 27 -38 . DOI: 10.11978/2020055
表1 菌株信息Tab. 1 Strain information |
| 物种名称 | 菌株 | 生物样品序列号 | 生物项目序列号 | 基因组编号 | 纯培养 | 呼吸类型 | 参考文献 |
|---|---|---|---|---|---|---|---|
| Caminibacter mediatlanticus | TB-2 | SAMN10884411 | PRJNA521379 | GCA_006459125.1 | 是 | 厌氧 | Giovannelli 等(2011) |
| Cetia pacifica | TB6 | SAMN03737945 | PRJNA284962 | GCA_003346815.1 | 是 | 厌氧 | Grosche等(2015) |
| Hydrogenimonas sp. | MAG | SAMN00210774 | PRJNA543187 | GCA_005886055.1 | 否 | 兼性厌氧 | Takai等(2004) |
| Lebetimonas sp.# | JS138 | SAMN03737948 | PRJNA66817 | GCA_003660105.1 | 否 | 厌氧 | Meyer等(2014) |
| Nautilia profundicola | AmH | SAMN03737951 | PRJNA284966 | GCA_003544915.1 | 是 | 厌氧 | Smith等(2008) |
| Nautilia sp. | PV-1 | SAMN03737952 | PRJNA284967 | GCA_003346755.1 | 是 | 厌氧 | Smith等(2008) |
| Nitratiruptor sp. | SB155-2 | SAMN03737954 | PRJNA66821 | GCA_003544935.1 | 是 | 兼性厌氧 | Nakagawa等(2005) |
| Sulfurimonas autotrophica | DSM 16294 | SAMN03737955 | PRJNA66823 | GCA_003346775.1 | 是 | 好氧 | Inagaki等(2003) |
| Campylobacter jejuni* | 81-176 | SAMEA4030738 | PRJEB6403 | GCA_900637395.1 | 是 | 微好氧 | Snelling等(2005) |
注: #表示没有完全测序; *表示试验用模式菌株 |
表2 质粒相关信息Tab. 2 Plasmid information |
| 质粒名称 | 质粒描述 |
|---|---|
| pGEX-6P | 用于构建GST融合蛋白表达载体 |
| pKNT25 | 细菌双杂交T25融合蛋白表达载体 |
| pUT18C | 细菌双杂交T18(在N端)融合蛋白表达载体 |
| pCH363 | 细菌双杂交T18(在C端)融合蛋白表达载体 |
表3 Campylobacter jejuni 81-176相关基因信息Tab. 3 Campylobacter jejuni 81-176 related genetic information |
| 基因 | 基因编号 | 蛋白结构域示意图 |
|---|---|---|
| cheV | CJJ81176_0311 |  |
| cheW | CJJ81176_0309 |  |
| tlp1 | CJJ81176_1498 |  |
| tlp2 | CJJ81176_0180 |  |
| tlp3# | CJJ81176_1548 |  |
| tlp3# | CJJ81176_1549 |  |
| tlp4 | CJJ81176_0289 |  |
| tlp5* | CJJ81176_0271-4 | Pseudogene |
| tlp6 | CJJ81176_0473 |  |
| tlp7 | CJJ81176_0975 |  |
| tlp8 | CJJ81176_1128 |  |
| tlp9 | CJJ81176_1205 |  |
| tlp10 | CJJ81176_0046 |  |
| aer1 | CJJ81176_1204 |  |
| aer2 | CJJ81176_1206 |  |
注: #表示Tlp3出现了点突变, 但还具有完整的MA结构域; *表示Tlp5出现了3次点突变, 为假基因 |
表4 引物序列Tab.4 Primer sequence |
| 引物名称 | 序列(5’→ 3’) |
|---|---|
| pKNT25+cheV_F | CTCTAGAGGATCCCCGGGTAATGTTTGATGAAAATATCGT |
| pKNT25+cheV_R | GTCATTGAATTCGAGCTCGGTTACCCCTGTTCTTGAGATT |
| pKNT25+cheW_F | CAGGTCGACTCTAGAGGATCCCCGGGTAATGAGTAATGAAAAATTAGAGCAAATTTTGC |
| pKNT25+cheW_R | CTGCATGGTCATTGAATTCGAGCTCGGAAATTCGCGCTTAAGTAAAGCTTCTACTTTG |
| pUT18C+tlp1_F | ACTGCAGGTCGACTCTAGATGAAATTAAAAAGATGCTTTTGGCTT |
| pUT18C+tlp1_R | ATATCGATGAATTGCTCGAGTTAAAATCTTTTTTTACTCACATCTTCAAG |
| pUT18C+tlp4_F | ACGCCACTGCAGGTCGACTCTAGATCTCTCCCCACTTGCAGCTATCCAAACAGGT |
| pUT18C+tlp4_R | TAGTTATATCGATGAATTGCTCGAGTTAAAACCTTTTCTTCTTAACATCTTCTAA |
| pUT18C+tlp6_F | ACGCCACTGCAGGTCGACTCTAGATATGTTTGGAAGTAAAATAAACCATTCTGAT |
| pUT18C+tlp6_R | TAGTTATATCGATGAATTGCTCGAGTTAATGATCTGACTCATCAAGCATTTCTTT |
| pUT18C+tlp7_F | ACTGCAGGTCGACTCTAGATCAATTTATCGAAAAAACTCATAAGGCAGTT |
| pUT18C+tlp7_R | ATATCGATGAATTGCTCGAGTTAAATTTGAAATTGGTTAAGTTCGCTTTC |
| pUT18C+tlp8_F | ACGCCACTGCAGGTCGACTCTAGATATGTTTGGTGCTAAGAAAAATAATACTGAA |
| pUT18C+tlp8_R | TAGTTATATCGATGAATTGCTCGAGTTATGACATCGCTTTAGCAACTTCAGCAGAGCT |
| pUT18C+tlp9_F | ACGCCACTGCAGGTCGACTCTAGATTTAGCAAACATAGAAGTGACAGCAAGATCT |
| pUT18C+tlp9_R | TAGTTATATCGATGAATTGCTCGAGTTATATTTTTAATTTTGCTAAGATTTCAGC |
| pUT18C+tlp10_F | ACTGCAGGTCGACTCTAGATTCAAGTCATCAAAATTCACAAAAACTCAA |
| pUT18C+tlp10_R | ATATCGATGAATTGCTCGAGTTACTGAAAGCTACTTAATTTTTCGGAGAG |
| pUT18C+aer1_F | ACTGCAGGTCGACTCTAGATAAAGAAATAGTTTTGTCTGAAAATGCTTTA |
| pUT18C+aer1_R | ATATCGATGAATTGCTCGAGTTAATTATTTTCTTGTAAGTTAAAAATAAGTTCAT |
| pUT18C+aer2_F | ACTGCAGGTCGACTCTAGATTCAAGAGAAATTTTTTTACAAGAAGATAGT |
| pUT18C+aer2_R | ATATCGATGAATTGCTCGAGTTATTTAGCTTCTTGAAGAGAAAAGATTAGC |
| pUT18C+tlp9 (no CZB-like)_F | ACTGCAGGTCGACTCTAGATTTAGCAAACATAGAAGTGACAGCAAGATCT |
| pUT18C+tlp9 (no CZB-like)_R | ATATCGATGAATTGCTCGAGTTAATAAAGAATATGATCGATTTTAACCAC |
| pCH363+tlp9_F | ACAGCTATGACCATGATTACGCCATTAGCAAACATAGAAGTGACAGCAAGATCT |
| pCH363+tlp9_R | TGGCCTCGCTGGCGGCTGAATTCGATATTTTTAATTTTGCTAAGATTTCAGCACT |
| pGEX-6P+tlp6_F | TCTGTTCCAGGGGCCCCTGGGATCCTTGCTTAGACAACACAAAGATGAGC |
| pGEX-6P+tlp6_R | GCTCGAGTCGACCCGGGAATTCCGGTTATATTTTTAATTTTGCTAAGATTTCAGCAC |
| pGEX-6P+tlp9_F | TCTGTTCCAGGGGCCCCTGGGATCCATGTTTGGAAGTAAAATAAACCATTCTGATCTTC |
| pGEX-6P+tlp9_R | GCTCGAGTCGACCCGGGAATTCCGGTTAATGATCTGACTCATCAAGCATTTC |
| pGEX-6P+tlp9 (no CZB-like)_F | TCTGTTCCAGGGGCCCCTGGGATCCATGTTTGGAAGTAAAATAAACCATTCTGATCTTC |
| pGEX-6P+tlp9 (no CZB-like)_R | GCTCGAGTCGACCCGGGAATTCCGGTTAATCGATTTTAACCACAGACAAAATCAATC |
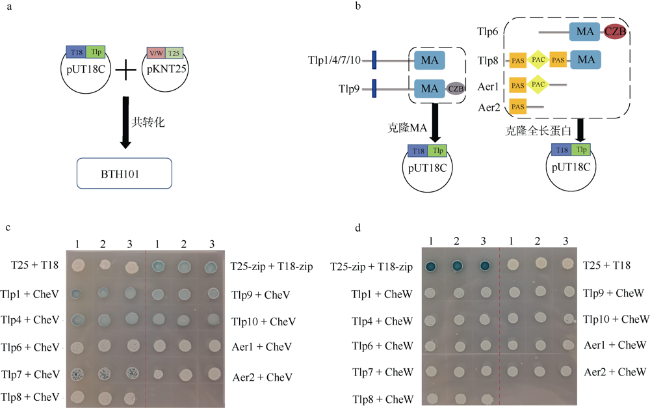
图2 细菌双杂交研究Campylobacter jejuni 81-176的CheV与趋化受体蛋白的相互作用a. 细菌双杂交操作示意图; b. 基因克隆示意图, 左图表示跨膜的Tlp只截取胞内的MA 结构域及MA 结构域后的序列进克隆, 右图表示胞内趋化受体选取蛋白的全长序列进行克隆; c. CheV与趋化受体蛋白的细菌双杂交结果; d. CheW与趋化受体蛋白的细菌双杂交结果。图c和图d中颜色深浅代表蛋白相互作用的强弱程度。BTH101为细菌双杂交系统菌株 Fig. 2 Using bacterial two-hybrid system to study the interaction between CheV and chemotactic receptors in Campylobacter jejuni 81-176 (a) Schematic diagram of bacterial two-hybrid experiment; (b) Schematic diagram of gene cloning. The left panel shows that for transmembrane Tlps, only MA domains and sequences after the MA domain were cloned into vectors. The right panel shows the full-length sequence of the cytolasmic chemotactic receptor was selected for cloning; (c) Interaction of CheV with Tlps by bacterial two-hybrid experiments; (d) Interaction of CheW with Tlps by bacterial two-hybrid experiments. The shade of color in Figure C and D represents the strength of protein interaction. BTH101 is a strain for bacterial double hybrid system |
表5 深海Epsilon-变形菌的F类型和CheV的分析统计Tab. 5 Analysis and statistics of F-Class and CheV in deep sea Epsilon-proteobacteria |
| 菌株 | F类型 | CheV蛋白编号 | 菌株 | F类型 | CheV蛋白编号 |
|---|---|---|---|---|---|
| Campylobacter jejuni* | 1 (F3) | WP_002857365.1 | Nautilia profundicola | 1 (F3) | WP_041361491.1 |
| Caminibacter mediatlanticus | 1 (F3) | WP_007474092.1 | Nautilia sp. PV-1 | 1 (F3) | WP_127679461.1 |
| Cetia pacifica | 1 (F3) | WP_123351600.1 | Nitratiruptor sp. SB155-2 | 1 (F14) | WP_012081979.1 |
| Hydrogenimonas sp. MAG | 1 (F3) | BBG65199.1 | Sulfurimonas autotrophica | 2 (F3/F8) | WP_013326429.1 WP_013327173.1# |
| Lebetimonas sp. JS138 | 1 (F3) | WP_024787070.1 |
注: *表示非深海菌株; #表示F8类型 |
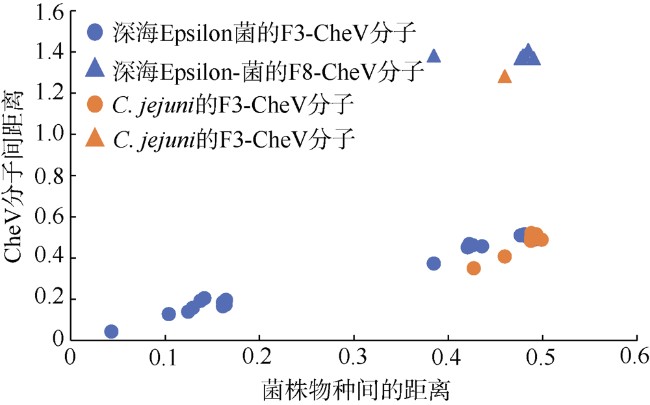
图3 深海Epsilon-变形菌和Campylobacter jejuni的CheV分子间进化距离分析蓝色表示深海Epsilon-变形菌的两两比较; 橙色表示深海Epsilon-变形菌与C. jejuni 81-176的两两比较; 圆圈表示不同菌株的F3-CheV间的距离; 三角形表示Sulfurimonas autotrophica的F8-CheV与其他Epsilon-变形菌菌株的F3-CheV之间的距离 Fig. 3 Pairwise Distances matrix to calculate the evolutionary distance for CheV homologs between deep-sea Epsilon-proteobacteria and Campylobacter jejuni Blue indicates the evolutionary distance between deep-sea Epsilon- proteobacteria. Orange indicates the evolutionary distance between deep-sea Epsilon-proteobacteria and C. jejuni 81-176. Dot shows the distance between F3-CheV in different Epsilon-proteobacteria, and triangle shows the distance between F8-CheV in sulfurimonas autotrophica and F3-CheV in other Epsilon-proteobacteria |
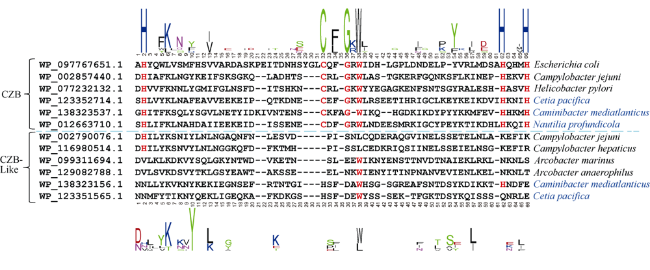
图4 CZB结构域和CZB-like结构域的序列比对分析通过多序列比对寻找保守氨基酸并以此区分CZB结构域和CZB-like结构域。蓝色字体代表的是深海菌株, 标红的字母代表CZB结构域的保守氨基酸位点 Fig. 4 Sequence alignment of CZB domain and CZB-like domain Blue font indicates deep-sea strain, Red letter indicates conserved amino acids of CZB domain |
表6 深海Epsilon-变形菌CZB结构域的分析统计Tab. 6 Analysis and statistics of CZB domain in deep sea Epsilon-proteobacteria |
| 菌株 | CZB个数 | 含CZB的蛋白编号 | CZB-like个数 | 含CZB-like的蛋白编号 |
|---|---|---|---|---|
| Campylobacter jejuni* | 1 | WP_002857440.1 | 1 | WP_002790076.1 |
| Caminibacter mediatlanticus | 2 | WP_138323537.1, WP_138323100.1 | 1 | WP_138323156.1 |
| Cetia pacifica | 2 | WP_123352714.1, WP_123352123.1 | 1 | WP_123351565.1 |
| Hydrogenimonas sp. MAG | 5 | BBG65387.1, BBG65955.1, BBG65459.1, BBG65926.1, BBG65424.1 | 0 | 0 |
| Lebetimonas sp. JS138 | 1 | WP_024791915.1 | 0 | 0 |
| Nautilia profundicola | 2 | WP_012663710.1, WP_015902667.1 | 0 | 0 |
| Nautilia sp. PV-1 | 2 | WP_127679641.1, WP_127679872.1 | 0 | 0 |
| Nitratiruptor sp. SB155-2 | 2 | WP_012081746.1, WP_012081977.1 | 0 | 0 |
| Sulfurimonas autotrophica | 3 | WP_013327874.1, WP_041675273.1, WP_013327332.1 | 0 | 0 |
注: *表示非深海菌株 |
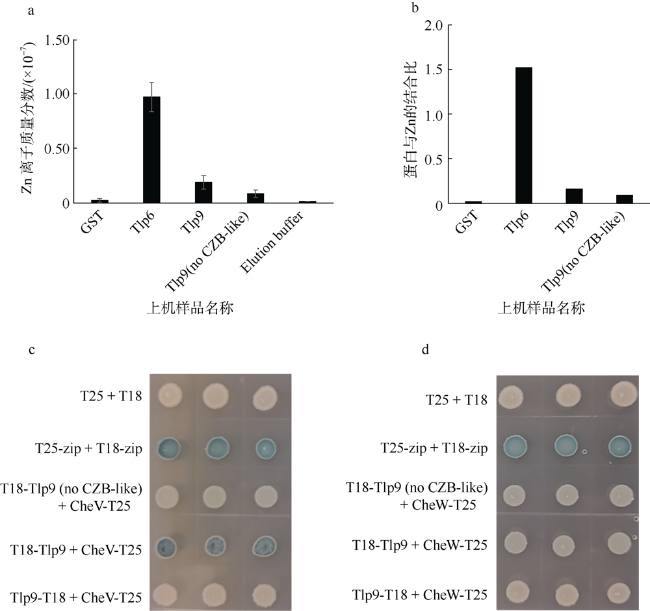
图5 CZB-like结构域的Zn结合能力及潜在功能的研究a. ICP-Mass检测CZB-like结构域的Zn离子结合能力; b. 根据3次质谱测定的平均离子浓度和平均蛋白浓度计算蛋白和Zn离子的结合比例; c和d. 细菌双杂交研究Tlp9的CZB-like结构域能否影响Tlp9与CheV和CheW的相互作用。图c和图d中不同颜色分别代表蛋白相互作用的强弱程度 Fig. 5 Study on Zn-binding capacity and potential function of CZB-like domain (a) Zn concentration measure by ICP-mass; (b) Using the average Zn concentration and average protein concentration, the binding ratio of protein and Zn is calculated; (c and d) To investigate whether the CZB-like domain of Tlp9 can affect the interaction with CheV or CheW by bacterial two-hybrid experiments. The shade of color in Figure C and D represents the strength of protein interaction |
| [1] |
王风平, 周悦恒, 张新旭, 等, 2013. 深海微生物多样性[J]. 生物多样性, 21(4):445-455.
|
| [2] |
张云怡, 曾令兵, 郭晓奎, 等, 2011. 细菌趋化过程中信号转导系统研究[J]. 中国微生态学杂志, 23(1):93-96 (in Chinese).
|
| [3] |
臧扬, 高贝乐, 2017. 深海热液口Epsilon-变形菌的物种多样性与环境适应机理[J]. 微生物学报, 57(9):1392-1399.
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}