
Journal of Tropical Oceanography >
Physiological and genomic characteristics of two lytic phages infecting Vibrio alginolyticus
Copy editor: YAO Yantao
Received date: 2020-06-24
Revised date: 2020-07-26
Online published: 2020-07-29
Supported by
Project of Program of Science and Technology of Guangzhou(20190420029)
National Natural Science Foundation of China(41506095)
National Natural Science Foundation of China(41676075)
Project of Innovation Academy of South China Sea Ecology and Environmental Engineering, Chinese Academy of Sciences(ISEE2019ZR02)
Project of Department of Science and Technology of Guangdong Province(2018B030320005)
China Postdoctoral Science Foundation(2019M662237)
Copyright
Phage therapy of bacterial diseases was considered to be effective and environmentally friendly in aquaculture. To obtain the biological agents that prevent Vibro alginolyticus, two jumbo phages vB_ValM_R10Z and vB_ValM_R11Z were isolated from prawn farming waters and the sewage in fishery markets in Dongshan Island, Fujian Province, using Vibrio alginolyticus ATCC 17749 T as the host. The morphology, host range, life cycle, and genome were identified and analyzed. Phages R10Z and R11Z had similar morphology of plaques and virions, and shared the same host range of infecting other V. owensii strain JL3186. Both phages were not sensitive to chloroform, indicating that the capsid did not contain lipids. They had the same latent period of 20 min but different blast sizes of 45 PFU·cell -1 for R10Z and 114 PFU·cell-1 for R11Z. The genomic sizes of phages R10Z and R11Z were 247167 bp and 246831 bp with G+C content of 41.30% and 41.33%, respectively. Furthermore, their genomes had a similarity of 99.45%, without virulence genes and antibiotic resistance genes. Our phylogenetic analysis showed that they belonged to genus Schizotequatrovirus, subfamily Tevenvirinae, family Myovirdae. Their homologous genes were distributed in the environment widely. Phages R10Z and R11Z had good capability and great potentials for their usage as phage therapy agents of vibriosis in aquaculture.
Key words: Vibrio; phage; phage therapy; physiological properties; genome
ZHONG Wanxuan , YANG Yunlan , LI Xiangfu , XU Jie . Physiological and genomic characteristics of two lytic phages infecting Vibrio alginolyticus[J]. Journal of Tropical Oceanography, 2021 , 40(4) : 22 -34 . DOI: 10.11978/2020067
图1 噬菌体vB_ValM_R10Z和vB_ValM_R11Z的噬菌斑形态图与电镜形态图a. vB_ValM_R10Z的噬菌斑形态图; b. vB_ValM_R11Z的噬菌斑形态图; c. vB_ValM_R10Z的电镜形态图; d. vB_ValM_R11Z的电镜形态图 Fig. 1 Phage plaques on double-layer plate and morphology in transmission electron microscopy of phages vB_ValM_R10Z and vB_ValM_R11Z a. Phage plaques of vB_ValM_R10Z; b. phage plaques of vB_ValM_R11Z; c. transmission electron microscopy images of vB_ValM_R10Z; d. transmission electron microscopy images of vB_ValM_R11Z |
表1 噬菌体vB_ValM_R10Z和vB_ValM_R11Z的裂解谱Tab. 1 The host ranges of vB_ValM_R10Z and vB_ValM_R11Z |
| 细菌种属 | 株系 | 致病性 | vB_ValM_R10Z | vB_ValM_R11Z |
|---|---|---|---|---|
| V. alginolyticus | ATCC 17749T | 可致病 | + | + |
| V. alginolyticus | JL2674 | 未知 | - | - |
| V. campbellii | JL2671 | 未知 | - | - |
| V. campbellii | JL3506 | 未知 | - | - |
| V. parahemolyticus | ATCC 17802T | 可致病 | - | - |
| V. parahemolyticus | BVP2 | 未知 | - | - |
| V. parahemolyticus | 20160623-13 | 可致病 | - | - |
| V. parahemolyticus | 20160719-2 | 可致病 | - | - |
| V. parahemolyticus | 20160615-5 | 可致病 | - | - |
| V. harveyi | BVH1 | 可致病 | - | - |
| V. harveyi | 20150916-2 | 可致病 | - | - |
| V. owensii | JL2581 | 未知 | - | - |
| V. owensii | JL3186 | 未知 | + | + |
| V. owensii | JL3187 | 未知 | - | - |
| V. caribbeanicus | JL3219 | 未知 | - | - |
| V. fortis | JL3515 | 未知 | - | - |
| V. chagasii | JL3518 | 未知 | - | - |
| V. inhibens | JL3707 | 未知 | - | - |
| V. cholerae | 20161020-5 | 可致病 | - | - |
| V. variabilis | JL3833 | 未知 | - | - |
注: “+”表示噬菌体能够侵染该细菌; “-”表示噬菌体不可侵染该细菌 |
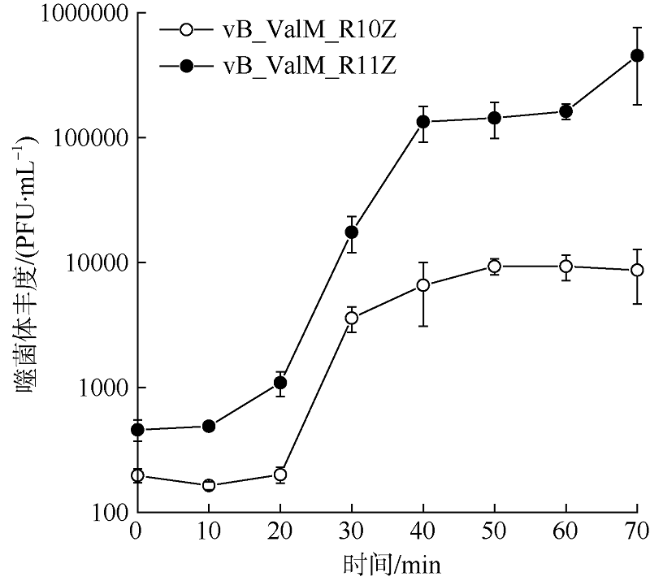
图2 噬菌体vB_ValM_R10Z和vB_ValM_R11Z的一步生长曲线图两株噬菌体分别进行生长曲线实验, 同时测试3个平行样品并取平均值, 误差线表示3个平行样品的标准差 Fig. 2 One-step growth curves of phages vB_ValM_R10Z and vB_ValM_R11Z Two phages were tested separately. Three parallel samples were tested at the same time, and the mean value was taken. The error bar represents the standard deviation of the parallel samples |
图3 Schizotequatrovirus(属)噬菌体的比较基因组分析本图采用Easyfig 2.2.3绘制; 红色箭头表示ORF, 深灰色连线表示相似度较高, 浅灰色连线表示相似度较低, 白色表示不相似 Fig. 3 Genomic comparison of genus Schizotequatrovirus Drawn with Easy |
表2 噬菌体vB_ValM_R10Z和vB_ValM_R11Z的tRNA基因Tab. 2 tRNA genes of phages vB_ValM_R10Z and vB_ValM_R11Z |
| vB_ValM_R10Z | vB_ValM_R11Z | |||||||
|---|---|---|---|---|---|---|---|---|
| tRNA基因 序号 | 位置 | 氨基酸 | 反密码子 | tRNA基因 序号 | 位置 | 氨基酸 | 反密码子 | |
| 1 | 164151~164264 | Phe | AAA | 1 | 163682~163794 | Phe | AAA | |
| 2 | 164275~164399 | Asn | GTT | 2 | 163805~163929 | Asn | GTT | |
| 3 | 164784~164861 | Met | CAT | 3 | 164314~164391 | Met | CAT | |
| 4 | 166706~166779 | Pro | TGG | 4 | 166236~166309 | Pro | TGG | |
| 5 | 166789~166865 | Pro | TGG | 5 | 166319~166395 | Pro | TGG | |
| 6 | 166873~166948 | Arg | ACG | 6 | 166404~166479 | Arg | ACG | |
| 7 | 167302~167378 | Leu | TAG | 7 | 166833~166909 | Leu | TAG | |
| 8 | 167380~167454 | Phe | GAA | 8 | 166911~166985 | Phe | GAA | |
| 9 | 167544~167618 | Lys | TTT | 9 | 167075~167149 | Lys | TTT | |
| 10 | 167630~167706 | Lys | TTT | 10 | 167161~167237 | Lys | TTT | |
| 11 | 167770~167844 | Glu | TTC | 11 | 167301~167375 | Glu | TTC | |
| 12 | 167851~167936 | Tyr | GTA | 12 | 167382~167467 | Tyr | GTA | |
| 13 | 168039~168115 | Met | CAT | 13 | 167570~167646 | Met | CAT | |
| 14 | 168411~168509 | Ser | TGA | 14 | 167942~168040 | Ser | TGA | |
| 15 | 168574~168647 | Ile | GAT | 15 | 168105~168178 | Ile | GAT | |
| 16 | 168654~168727 | Trp | CCA | 16 | 168185~168258 | Trp | CCA | |
| 17 | 168803~168877 | Asn | GTT | 17 | 168334~168408 | Asn | GTT | |
| 18 | 168879~168954 | Asn | GTT | 18 | 168410~168485 | Asn | GTT | |
| 19 | 169024~169099 | Asp | GTC | 19 | 168554~168629 | Asp | GTC | |
| 20 | 169119~169192 | Gln | TTG | 20 | 168649~168722 | Gln | TTG | |
| 21 | 169322~169396 | Thr | TGT | 21 | 168850~168925 | Thr | TGT | |
| 22 | 169968~170042 | Ala | TGC | 22 | 169167~169241 | Ala | TGC | |
| 23 | 170117~170192 | Arg | TCT | 23 | 169316~169391 | Arg | TCT | |
| vB_ValM_R10Z | vB_ValM_R11Z | |||||||
| tRNA基因 序号 | 位置 | 氨基酸 | 反密码子 | tRNA基因 序号 | 位置 | 氨基酸 | 反密码子 | |
| 24 | 170717~170792 | Met | CAT | 24 | 169915~169990 | Met | CAT | |
| 25 | 170982~171055 | Gly | TCC | 25 | 170180~170253 | Gly | TCC | |
| 26 | 171062~171135 | Val | TAC | 26 | 170260~170333 | Val | TAC | |
| 27 | 171551~171627 | His | GTG | 27 | 170749~170825 | His | GTG | |
| 28 | 171640~171713 | Cys | GCA | 28 | 170838~-170911 | Cys | GCA | |
| 29 | 171716~171792 | Leu | TAA | 29 | 170914~170990 | Leu | TAA | |
| 30 | 171909~171985 | Leu | CAA | 30 | 171107~171183 | Leu | CAA | |
| 31 | 172154~172242 | Ser | GCT | 31 | 171352~171440 | Ser | GCT | |
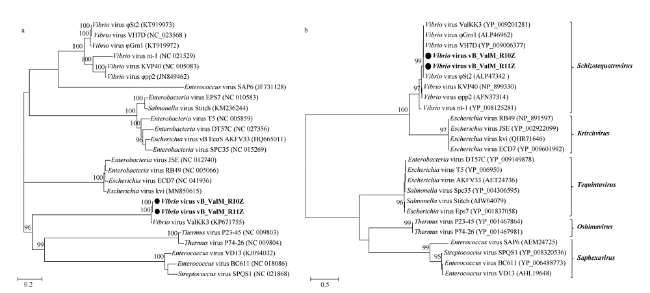
图4 噬菌体全基因组(a)和DNA聚合酶基因(b)的系统发育进化树系统发育进化树由最大似然法构建, Bootstrap 检验1000次,检验可信度低于90的值隐藏; vB_ValM_R10Z 和vB_ValM_ RI0Z 加粗显示,括号内为噬菌体的GenBank序列号。系统发育树中包括本文分离获得的噬菌体vB__ValM_R10Z、vB__ValM_R1IZ和所有前人已发表的Schizotequatrovirus类噬菌体,以及其余类群的部分代表性噬菌体 Fig. 4 Phy logenetic trees of complete genome (a) and DNA polymerase (b) of phages vB_ ValM_ R10Z, vB_ ValM_R11Z and other known phages Maximum likelihood analyses with 1000 bootstrap replicates were used to derive the trees based on amino-acid sequences. Bootstrap values are shown above the major nodes, and the values below 90 are hidden. Phages vB_ValM_R10Z and vB_ValM_R11Z are marked in bold. Numbers in brackets represent corresponding GenBank ID. The phylogenetic trees include phages vB_ValM_ R10Z and vB_ValM_ R11Z, as well as all published Schizotequatrovirus phages and some representative phages of other genera |
表3 Schizotequatrovirus噬菌体的对比Tab. 3 Comparison of Schizotequatrovirus phages |
| 噬菌体 | vB_ValM_R10Z | vB_ValM_R11Z | VH7D | φSt2 | φGrn1 | ValKK3 | KVP40 | φpp2 | nt-1 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 宿主菌 | V. alginolyticus | V. alginolyticus | V. sp. 7D | V. alginolyticus | V. alginolyticus | V. alginolyticus | V. parahaemolyticus | V. parahaemolyticus | V. natrigens | ||||||||||
| 地点 | 中国东山岛 | 中国东山岛 | 中国厦门 | 希腊克里特岛 | 希腊克里特岛 | 马来西亚 | 日本Urado湾 | 中国台湾南部 | 大西洋 | ||||||||||
| 分离环境 | 养殖水体 | 污水 | 养殖水体 | 海水 | 海水 | 沉积物 | 污水 | 养殖水体 | 沉积物 | ||||||||||
| 头长/nm | 147 | 148 | 155 | 151 | 138 | 140 | 140 | 150 | 120 | ||||||||||
| 头宽/nm | 78 | 81 | 75 | 81 | 74 | 79 | 70 | 90 | 70 | ||||||||||
| 尾长/nm | 120 | 114 | 132 | 134 | 120 | 130 | 110 | 110 | |||||||||||
| 潜伏期/min | 20 | 20 | 30 | 30 | |||||||||||||||
| 裂解量/PFU·cell-1 | 43 | 114 | 97 | 44 | |||||||||||||||
| 噬菌体 | vB_ValM_R10Z | vB_ValM_R11Z | VH7D | φSt2 | φGrn1 | ValKK3 | KVP40 | φpp2 | nt-1 | ||||||||||
| GenBank 收录号 | MT612988 | MT612989 | KC131129.1 | KT919973.1 | KT919972 | KP671755.1 | AY283928 | JN849462 | HQ317393 | ||||||||||
| 基因组大小/bp | 247167 | 246831 | 246964 | 250485 | 248605 | 248088 | 244835 | 246421 | 247489 | ||||||||||
| G+C值 | 41.30% | 41.33% | 41.31% | 42.60% | 38.80% | 41.20% | 42.60% | 42.55% | 41.33% | ||||||||||
| ORF数量/个 | 386 | 385 | 387 | 412 | 410 | 390 | 386 | 383 | 379 | ||||||||||
| tRNA数量/个 | 31 | 31 | 27 | 29 | 28 | 30 | 30 | 28 | |||||||||||
| 毒力基因 | 无 | 无 | 无 | 无 | 无 | 无 | 无 | 无 | 无 | ||||||||||
| 抗药性基因 | 无 | 无 | 无 | 无 | 无 | 无 | 无 | 无 | 无 | ||||||||||
| 参考文献 | 本文 | 本文 | Luo et al, 2015 | Kalatzis et al, 2016; Skliros et al, 2016 | Kalatzis et al, 2016; Skliros et al, 2016 | Lal et al, 2016 | Matsuzaki et al, 1992; Miller et al, 2003 | Lin et al, 2012 | Zachary, 1976 |
注: 表中空白表示无数据 |
| [1] |
冯书章, 刘军, 孙洋, 2007. 细菌的病毒——噬菌体最新分类与命名[J]. 中国兽医学报, 27(4):604-608 (in Chinese).
|
| [2] |
黄瑞芳, 2005. 斜带石斑鱼溶藻弧菌病的研究[J]. 水产科学, 24(6):1-3.
|
| [3] |
李自强, 2018. 哈维氏弧菌噬菌体的分离及其生物学特性[D]. 厦门: 厦门大学: 41-42.
|
| [4] |
刘淇, 王学忠, 戴芳钰, 等, 2007. 梭子蟹溶藻弧菌病的初步研究[J]. 齐鲁渔业, 24(9):1-3.
|
| [5] |
农业农村部渔业渔政管理局, 全国水产技术推广总站, 中国水产学会, 2019. 中国渔业统计年鉴-2019[M]. 北京: 中国农业出版社: 130-132.
MARA Bureau of Fisheries, National Fisheries Technology Extension Center, China Society of Fisheries, 2019. 2019 China fishery statistical yearbook[M]. Beijing: China Agriculture Press: 130-132(in Chinese).
|
| [6] |
张晓华, 林禾雨, 孙浩, 2018. 弧菌科分类学研究进展[J]. 中国海洋大学学报, 48(8):43-56.
|
| [7] |
张昕, 蔡俊鹏, 2004. 噬菌体在水产养殖中的应用[J]. 粮食与饲料工业, (6):38-39.
|
| [8] |
张正福, 纪来升, 1981. 密度梯度离心技术在分子生物学研究中的应用[J]. 植物生理学通讯, (6):60-64.
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}