
Journal of Tropical Oceanography >
The effect of high temperature on stress physiology of Sargassum fusiforme
Copy editor: YIN Bo
Received date: 2020-05-22
Request revised date: 2020-07-29
Online published: 2020-08-02
Supported by
Science and Technology Plan Projects in Dongtou District of Wenzhou City(N2016Y20B)
Major Fisheries and Agriculture Science and Technology Projects in Dongtou District of Wenzhou City(N2018Y03A)
Special program for basic business of Heilongjiang Provincial Department of Education(135209265)
Fundamental Research funds in Heilongjiang Provincial Universities (Special specialty of plant food processing technology)(YSTSXK201877)
Copyright
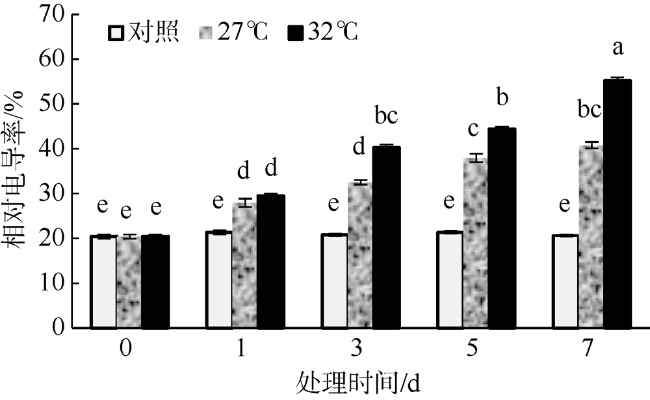
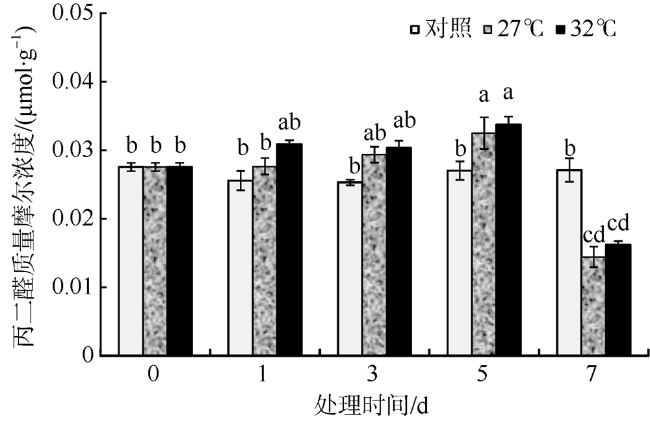
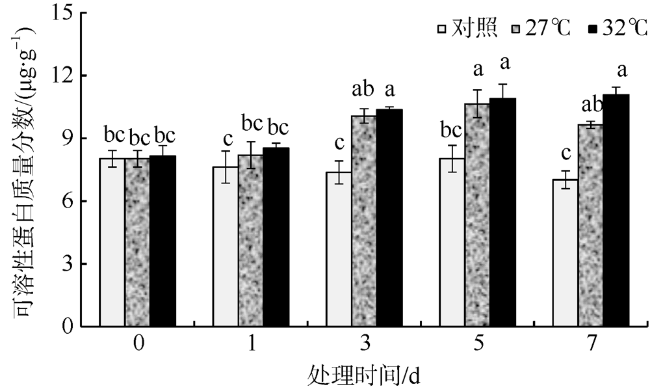
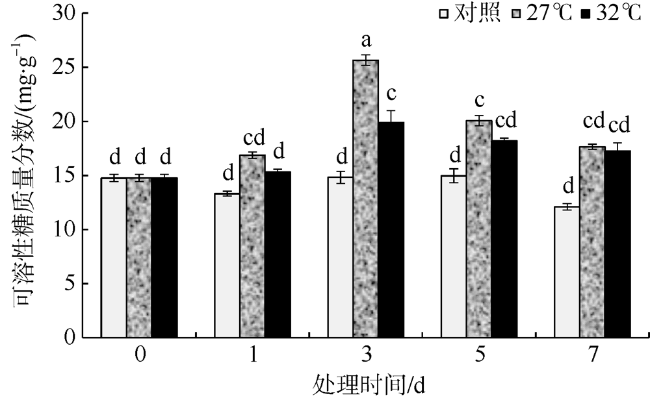
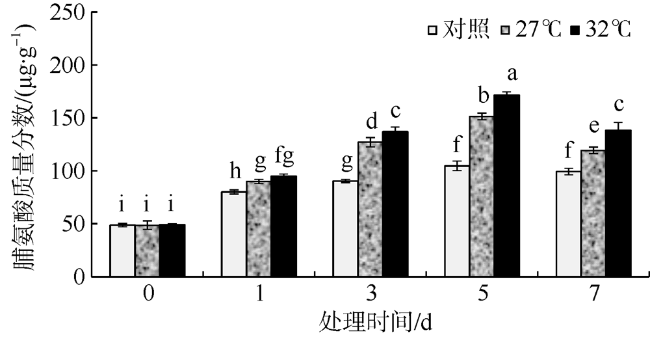
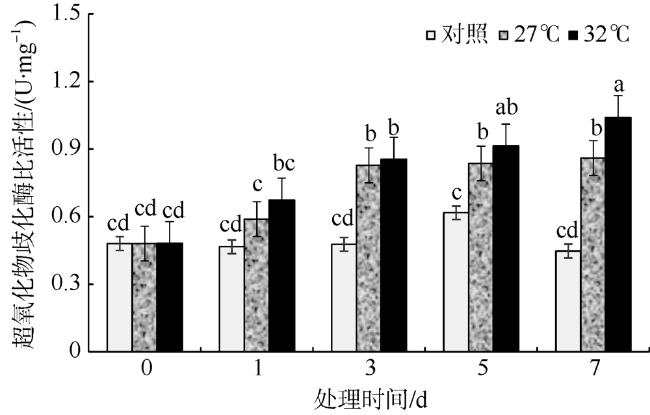
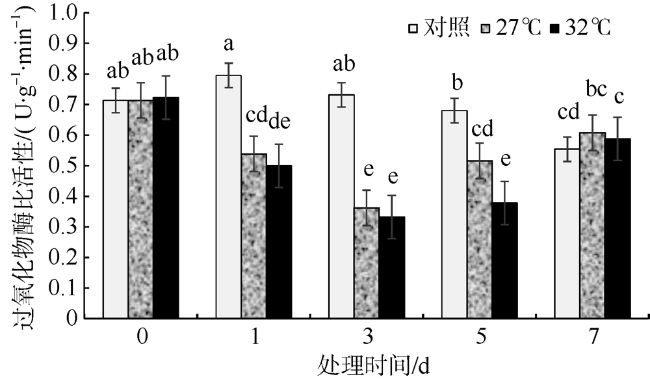
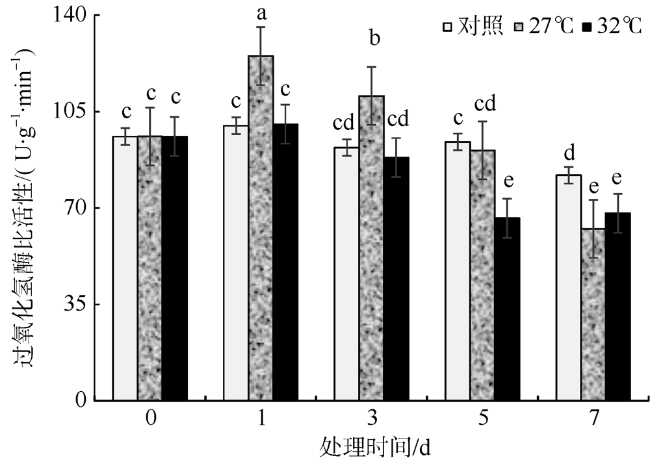
In this study, Sargassum fusiforme was treated at 27 ℃ and 32 ℃, and Sargassum fusiforme grown at 22 ℃ was used as control. Air bladders were collected at 0, 1, 3, 5, and 7 days of the treatment, respectively, to explore the effects of high temperature stress on membrane damage, osmotic regulation and antioxidant protection of Sargassum fusiforme. The results showed that the membrane system of Sargassum fusiforme was damaged by high temperature stress, which resulted in ion extravasation in vivo, and membrane lipid peroxidation; and the relative conductivity and Malondialdehyde (MDA) content of cells increased. Soluble protein, soluble sugar and proline were accumulated in the cells. Soluble protein was always higher than the control under high temperature stress, and reached the maximum value at 7 days. Soluble sugar and proline increased first and then decreased in the high temperature stress group, but they were always higher than those in the control group. Soluble sugar reached the maximum value at 3 days under high temperature stress, and proline reached the maximum value at 5 days under high temperature stress. After high temperature stress, Sargassum fusiforme can reduce the damage of high temperature stress mainly by increasing the activity of antioxidant enzyme Superoxide dismutase (SOD), while the activities of Peroxidase (POD) and Catalase (CAT) decreased in the process of high temperature stress. In a word, Sargassum fusiforme is more sensitive to high temperature stress. The higher the temperature is, the longer the treatment time is, the greater the damage to Sargassum fusiforme is.
LIU Lijie , LIN Lidong . The effect of high temperature on stress physiology of Sargassum fusiforme[J]. Journal of Tropical Oceanography, 2021 , 40(2) : 74 -82 . DOI: 10.11978/2020052
表1 试验期间的环境条件Tab. 1 The environment factors during experiments |
| 影响因素 | 光周期/(光照/黑暗) | 光强/(μmol·m-2·s-1) | 盐度/‰ | CO2质量浓度/(mg·m-3) | pH |
|---|---|---|---|---|---|
| 数值 | 12 : 12 | 151~155 | 35 | 900 | 7.8 |
| [1] |
苍晶, 赵会杰, 2013. 植物生理学实验教程[M]. 北京: 高等教育出版社: 141-142.
|
| [2] |
付峰, 隋正红, 李秉钧, 等, 2017. 热锻炼对龙须菜高温胁迫响应的影响[J]. 海洋科学, 41(4):10-16.
|
| [3] |
贺亮, 2017. 高温胁迫下半叶马尾藻中国变种生理生化响应及其耐热机制的初步研究[D]. 湛江: 广东海洋大学.
|
| [4] |
李合生, 2000. 植物生理生化实验原理和技术[M]. 2版. 北京: 高等教育出版社: 297-298.
|
| [5] |
李荣华, 郭培国, 张华, 等, 2012. 高温胁迫对不同耐热性菜心材料生理特性的差异研究[J]. 北方园艺, (1):1-6.
|
| [6] |
禄鑫, 2012. 高温胁迫对报春花叶片细胞膜透性的影响[J]. 天津农业科学, 18(2):140-141.
|
| [7] |
孙建璋, 方家仲, 朱植丰, 等, 1996. 羊栖菜Sargassum fusiforme (Harve) Setch繁殖生物学的初步研究[J]. 浙江水产学报, 15(4):243-249.
|
| [8] |
孙雪, 蔡西栗, 徐年军, 2013. 海洋红藻龙须菜对2种逆境温度胁迫的应激生理响应[J]. 水生生物学报, 37(3):535-540.
|
| [9] |
孙圆圆, 孙庆海, 孙建璋, 2009. 温度对羊栖菜生长的影响[J]. 浙江海洋学院学报(自然科学版), 28(3):342-347.
|
| [10] |
王凯红, 刘向平, 张乐华, 等, 2011. 5种杜鹃幼苗对高温胁迫的生理生化响应及耐热性综合评价[J]. 植物资源与环境学报, 20(3):29-35.
|
| [11] |
杨华庚, 颜速亮, 陈慧娟, 等, 2011. 高温胁迫下外源茉莉酸甲酯、钙和水杨酸对蝴蝶兰幼苗耐热性的影响[J]. 中国农学通报, 27(28):150-157.
|
| [12] |
曾呈奎, 陆保仁, 2000. 中国海藻志第三卷褐藻门第二册墨角藻目[M]. 北京: 科学出版社: 100.
|
| [13] |
张展, 刘建国, 刘吉东, 2002. 羊栖菜的研究述评[J]. 海洋水产研究, 23(3):67-74.
|
| [14] |
张治安, 陈展宇, 2008. 植物生理学实验技术[M]. 长春: 吉林大学出版社: 192-193.
|
| [15] |
张志良, 瞿伟菁, 李小方, 2015. 植物生理学实验指导[M]. 4版. 北京: 高等教育出版社: 125.
|
| [16] |
赵素芬, 姚伟立, 郭学中, 等, 2015. 温度与光照强度对羊栖菜幼孢子体生长率的联合效应[J]. 水产养殖, 36(10):42-47.
|
| [17] |
赵昕, 李玉霖, 2001. 高温胁迫下冷地型草坪草几项生理指标的变化特征[J]. 草业学报, 10(4):85-91.
|
| [18] |
赵勇竣, 徐术菁, 王钊, 2019. 高温胁迫对3个番茄品种生长和生理指标的影响[J]. 江苏农业科学, 47(17):147-149.
|
| [19] |
朱静, 杨再强, 李永秀, 等, 2012. 高温胁迫对设施番茄和黄瓜光合特性及抗氧化酶活性的影响[J]. 北方园艺, (1):63-68.
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}