
Journal of Tropical Oceanography >
Vertical distributions of epipelagic and mesopelagic zooplankton in the continental slope of the northeastern South China Sea*
Copy editor: YIN Bo
Received date: 2020-06-13
Request revised date: 2020-08-02
Online published: 2020-08-09
Supported by
National Natural Science Foundation of China(31971432)
National Natural Science Foundation of China(41976112)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0401)
Science and Technology Basic Resources Investigation Program of China(2017FY201404)
Strategic Priority Research Program of the Chinese Academy of Sciences(XDA11020200)
Copyright
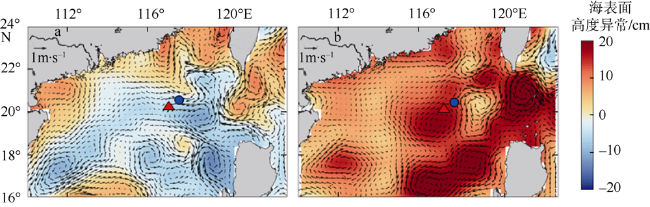
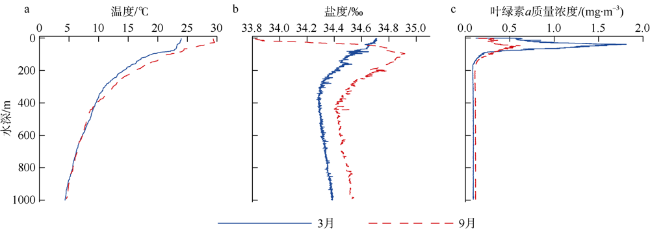
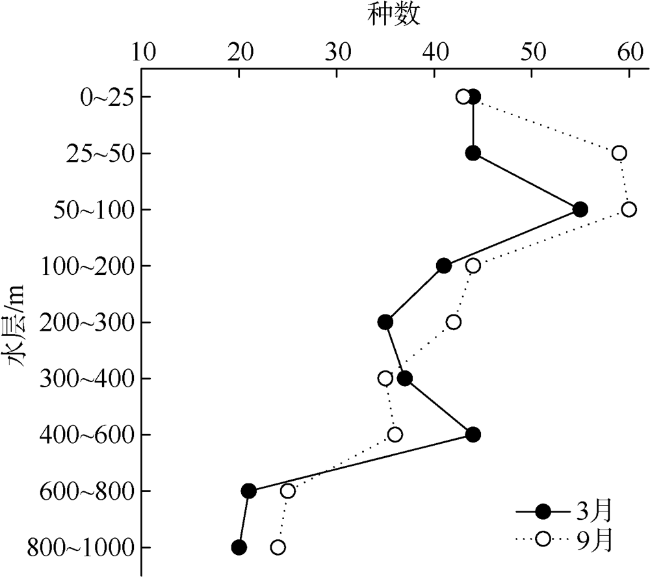
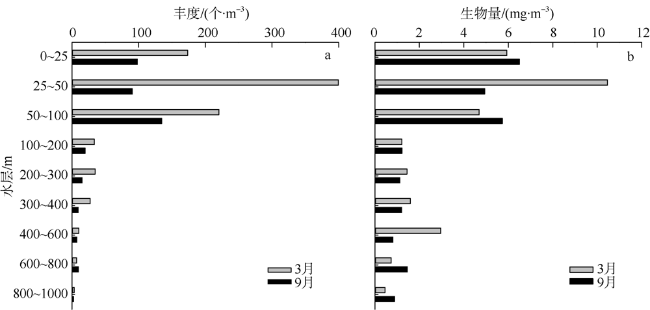
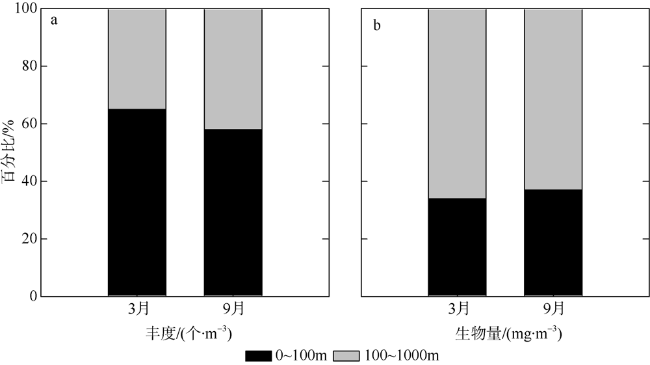
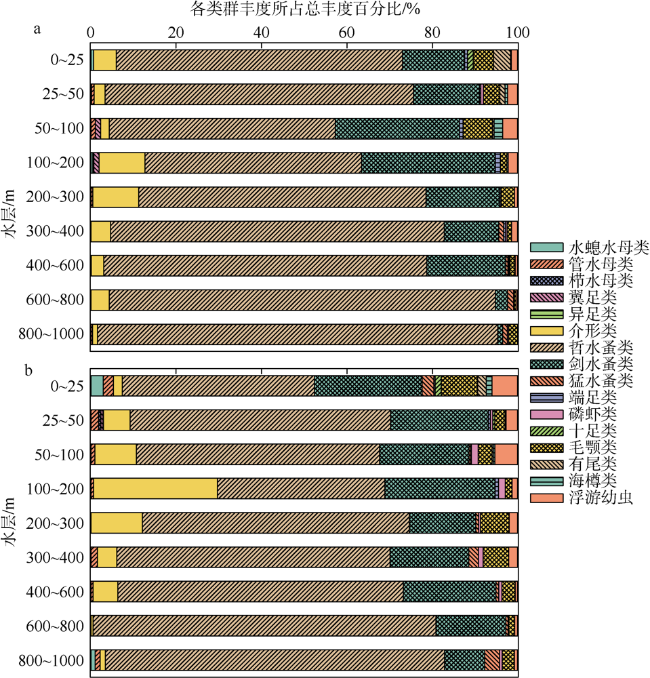
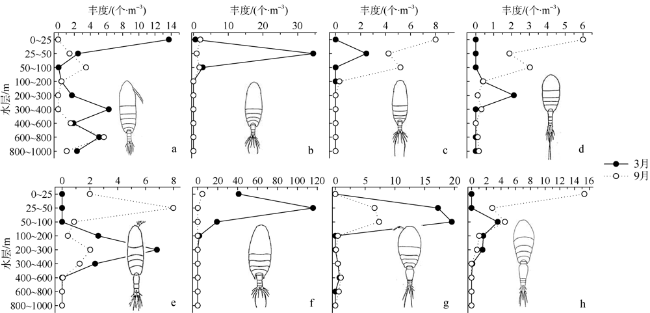
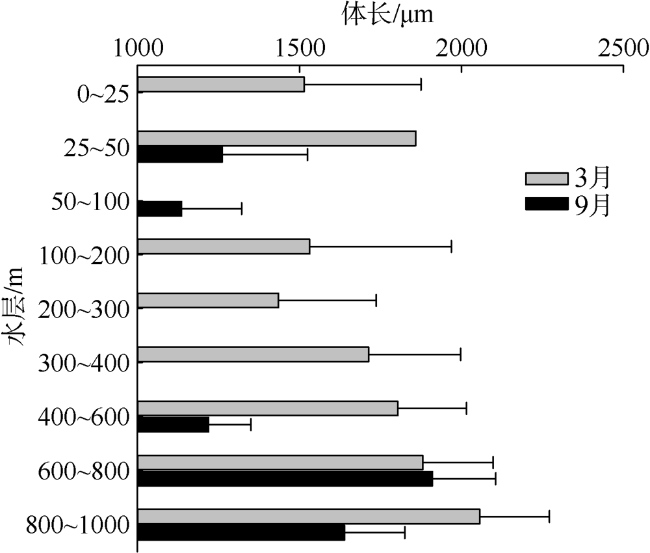
Based on the vertically stratified sampling of zooplankton in the continental slope of the northeastern South China Sea (SCS) in March and September 2016, the seasonal and vertical variations in zooplankton species composition, abundance, and biomass were analyzed and compared to explore the factors affecting their differences. A total of 225 species of zooplankton was identified; 150 species appeared in March, and 169 species, in September. There were 132 species of copepods, followed by 18 species of chaetognaths and siphonophores, respectively. The number of zooplankton species was the highest in the 50 ~ 100 m water layer and generally decreased with the increase of water depth. The abundance and biomass of zooplankton were mainly concentrated in the range of 0 ~ 100 m, and both of them decreased with the increase of depth in the deeper water layer. However, above 60% of the biomass concentrated within the layers between 100 and 1000 m. The abundance of dominant species had obvious seasonal and vertical variation. In March, the abundance of coastal species such as Undinula vulgaris and Canthocalanus pauper was higher above the depth of 100 m. In September, the abundance of oceanic species such as Cosmocalanus darwinii and Lucicutia flavicornis was high in the upper layer of 100 m. Calanoides carinatus showed higher abundance and average body length in March than in September, and the larger body length appeared mainly in deep layers. The zooplankton community could be divided into three groups of 0 ~ 100 m, 100 ~ 400 m, and 400 ~ 1000 m, which was mainly caused by the differences in the composition and abundance of different water layers. Zooplankton abundance and biomass were significantly positively correlated with temperature and chlorophyll a concentration. The seasonal and vertical variations in zooplankton abundance and biomass were driven by monsoon, coastal currents, and mesoscale eddies in the continental slope of the northeastern SCS.
LI Kaizhi , REN Yuzheng , KE Zhixin , LI Gang , TAN Yehui . Vertical distributions of epipelagic and mesopelagic zooplankton in the continental slope of the northeastern South China Sea*[J]. Journal of Tropical Oceanography, 2021 , 40(2) : 61 -73 . DOI: 10.11978/2020061
表1 南海东北部陆坡区浮游动物各类群在3月和9月出现的种数及所占百分比Tab. 1 Species richness and percentage of zooplankton groups in the continental slope of the northeastern South China Sea in March and September |
| 类群 | 3月种数 | 9月种数 | 合计 | 百分比/% |
|---|---|---|---|---|
| 水螅水母类Hydromedusae | 2 | 4 | 4 | 1.78 |
| 管水母类Siphonophores | 7 | 15 | 18 | 8.00 |
| 栉水母类Ctenophores | 0 | 1 | 1 | 0.44 |
| 浮游翼足类Pteropods | 3 | 2 | 5 | 2.22 |
| 浮游异足类Heteropods | 0 | 1 | 1 | 0.44 |
| 介形类Ostracods | 10 | 12 | 17 | 7.56 |
| 哲水蚤类Calanoids | 75 | 69 | 97 | 43.11 |
| 剑水蚤类Cyclopoids | 24 | 22 | 31 | 13.78 |
| 猛水蚤类Harpacticoids | 2 | 4 | 4 | 1.78 |
| 端足类Amphipods | 2 | 5 | 7 | 3.11 |
| 磷虾类Euphausiacea | 3 | 7 | 7 | 3.11 |
| 十足类Decapods | 1 | 2 | 2 | 0.89 |
| 毛颚类Chaetognaths | 12 | 15 | 18 | 8.00 |
| 有尾类Appendicularians | 2 | 2 | 2 | 0.89 |
| 全肌目类Doliolids | 1 | 2 | 2 | 0.89 |
| 半肌目类Salps | 1 | 1 | 2 | 0.89 |
| 火体虫类Pyrosomatids | 1 | 0 | 1 | 0.44 |
| 浮游幼虫Meroplankton | 4 | 5 | 6 | 2.67 |
| 合计 | 150 | 169 | 225 | 100 |
表2 南海东北部陆坡区3月和9月浮游动物优势种及优势度Tab. 2 Dominant species and their dominance in the continental slope of the northeastern South China Sea in March and September |
| 优势种 | 3月优势度 | 9月优势度 |
|---|---|---|
| 隆线似哲水蚤Calanoides carinatus | 0.037 | 0.023 |
| 微刺哲水蚤Canthocalanus pauper | 0.018 | 0.004 |
| 达氏筛哲水蚤Cosmocalanus darwinii | 0.000 | 0.026 |
| 黄角光水蚤Lucicutia flavicornis | 0.001 | 0.028 |
| 痩乳点水蚤Pleuromamma gracilis | 0.004 | 0.029 |
| 普通波水蚤Undinula vulgaris | 0.087 | 0.003 |
| 中隆水蚤Oncaea media | 0.019 | 0.033 |
| 丽隆水蚤Oncaea venusta | 0.003 | 0.035 |
图7 南海东北部陆坡区3月和9月浮游动物优势种丰度的垂直分布a. 隆线似哲水蚤; b. 微刺哲水蚤; c. 达氏筛哲水蚤; d. 黄角光水蚤; e. 瘦乳点水蚤; f. 普通波水蚤; g. 中隆水蚤; h. 丽隆水蚤。各图内图形为该种的形态图 Fig. 7 Vertical distributions of abundance of dominant species in the continental slope of the northeastern South China Sea in March and September (a) Calanoides carinatus; (b) Canthocalanus paupe; (c) Cosmocalanus darwinii; (d) Lucicutia flavicorni; (e) Pleuromamma gracilis; (f) Undinula vulgaris; (g) Oncaea medi; (h) Oncaea venusta |
表3 浮游动物丰度和生物量与环境因子的相关性系数Tab. 3 Correlation coefficients between zooplankton abundance and biomass and environmental factors |
| 变量 | 丰度/(个·m-3) | 生物量/( mg·m-3) | ||
|---|---|---|---|---|
| 3月 | 9月 | 3月 | 9月 | |
| 温度/℃ | 0.855* | 0.867* | 0.841* | 0.909*** |
| 盐度/‰ | 0.834* | 0.302n.s. | 0.833* | 0.056n.s. |
| 叶绿素a质量浓度/(mg·m-3) | 0.877* | 0.757* | 0.931*** | 0.790* |
注: *表示p<0.05, 相关性显著; ***表示p<0.001, 相关性极显著; n.s.表示不相关 |
| [1] |
陈清潮, 章淑珍, 1965. 黄海和东海浮游桡足类Ⅰ: 哲水蚤目[J]. 海洋科学集刊, 7: 20-131.
|
| [2] |
陈清潮, 章淑珍, 朱长寿, 1974. 黄海和东海的浮游桡足类Ⅱ: 剑水蚤目和猛水蚤目[J]. 海洋科学集刊, 9: 27-100.
|
| [3] |
陈瑞祥, 蔡秉及, 林茂, 等, 1988. 南海中部海域浮游动物的垂直分布[J]. 海洋学报, 10(3):337-341.
|
| [4] |
程旭华, 齐义泉, 王卫强, 2005. 南海中尺度涡的季节和年际变化特征分析[J]. 热带海洋学报, 24(4):51-59.
|
| [5] |
杜飞雁, 王雪辉, 谷阳光, 等, 2014. 南沙群岛西南大陆斜坡海域浮游动物的垂直分布[J]. 海洋学报, 36(6):94-103.
|
| [6] |
冯士筰, 李凤岐, 李少菁, 1999. 海洋科学导论[M]. 北京: 高等教育出版社: 1-503.
|
| [7] |
龚玉艳, 杨玉滔, 范江涛, 等, 2017. 南海北部陆架斜坡海域夏季浮游动物群落的空间分布[J]. 南方水产科学, 13(5):8-15.
|
| [8] |
龚玉艳, 杨玉滔, 孔啸兰, 等, 2018. 南海北部陆坡海域瓦氏眶灯鱼的渔业生物学特征[J]. 中国水产科学, 25(5):1091-1101.
|
| [9] |
连光山, 钱宏林, 1984. 西太平洋热带水域的浮游桡足类[C]// 西太平洋热带水域浮游生物论文集. 北京: 海洋出版社: 118-205.
|
| [10] |
刘长建, 庄伟, 夏华永, 等, 2012. 2009—2010年冬季南海东北部中尺度过程观测[J]. 海洋学报, 34(1):8-16.
|
| [11] |
舒业强, 王强, 俎婷婷, 2018. 南海北部陆架陆坡流系研究进展[J]. 中国科学D辑: 地球科学, 48(3):276-287.
|
| [12] |
苏纪兰, 2005. 南海环流动力机制研究综述[J]. 海洋学报, 27(6):1-8.
|
| [13] |
孙栋, 王春生, 2017. 深远海浮游动物生态学研究进展[J]. 生态学报, 37(10):3219-3231.
|
| [14] |
王翠, 郭晓峰, 方婧, 等, 2018. 闽浙沿岸流扩展范围的季节特征及其对典型海湾的影响[J]. 应用海洋学学报, 37(1):1-8.
|
| [15] |
徐兆礼, 陈亚瞿, 1989. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系[J]. 生态学杂志, 8(4):13-15, 19.
|
| [16] |
尹健强, 黄良民, 李开枝, 等, 2013. 南海西北部陆架区沿岸流和上升流对中华哲水蚤分布的影响[J]. 海洋学报, 35(2):143-153.
|
| [17] |
于君, 邱永松, 2016. 黑潮入侵对南海东北部初级生产力的影响[J]. 南方水产科学, 12(4):17-27.
|
| [18] |
张博, 曾丽丽, 陈举, 等, 2018. 基于南海北部开放航次观测的2004—2005年次表层盐度异常特征与形成机制[J]. 海洋与湖沼, 49(1):9-16.
|
| [19] |
张谷贤, 陈清潮, 1983. 南海北部和中部的毛颚类[C]// 南海海洋生物研究论文集(一). 北京: 海洋出版社: 17-63.
|
| [20] |
张金标, 2005. 中国海洋浮游管水母类[M]. 北京: 海洋出版社: 1-151.
|
| [21] |
张武昌, 赵楠, 陶振铖, 等, 2010. 中国海洋浮游桡足类图谱[M]. 北京: 科学出版社: 1-468.
|
| [22] |
中国科学院南海海洋研究所, 1985. 南海海区综合调查研究报告(二)[M]. 北京: 科学出版社: 357-378(in Chinese).
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}