
Journal of Tropical Oceanography >
Preliminary study on immunity function of aquaporin AQP4 in the pearl oyster Pinctada fucata martensii
Copy editor: YIN Bo
Received date: 2020-05-22
Request revised date: 2020-08-16
Online published: 2020-08-31
Supported by
Earmarked Fund for Modern Agro-industry Technology Research System(CARS-49)
Innovation Academy of South China Sea Ecology and Environmental Engineering, Chinese Academy of Sciences(ISEE2018PY03)
Shellfish and Large Algae Industry Innovation Team Project of Guangdong Province, China(2019KJ146)
Science and Technology Planning Project of Guangdong Province, China(2017B0303014052)
Copyright
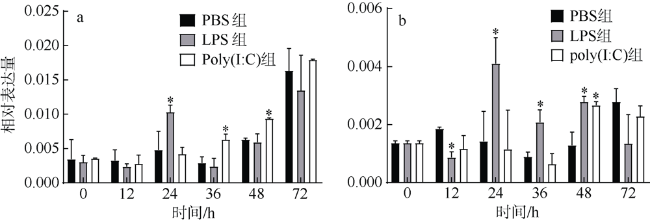
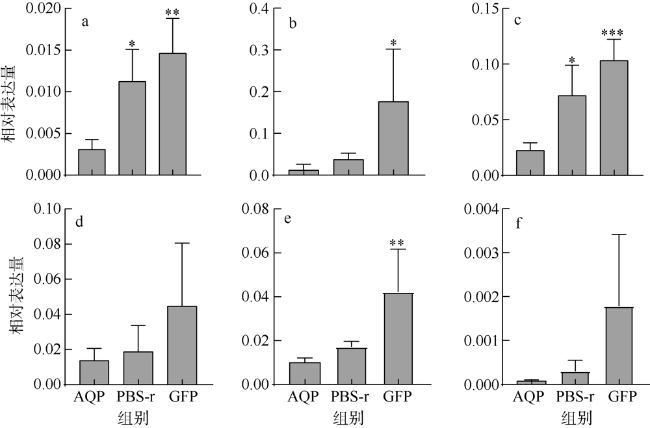
To identify the immunity function of Aquaporin of Pinctada fucata martensii (PfAQP4), the mRNA expression patterns of PfAQP4 and immune-related genes were detected by qPCR after immunostimulation and RNA interference. The results showed that the mRNA expression of PfAQP4 in the mantle was significantly up-regulated at 24 h (p < 0.05), while that in the digestive gland was significantly down-regulated at 12 h, and increased at 24 h, 36 h and 48 h after LPS injection (p < 0.05). After poly (I : C) injection, the expression of PfAQP4 was significantly up-regulated in the mantle at 36 h and 48 h (p < 0.05), and up-regulated at 48 h in the digestive gland (p < 0.05). When PfAQP4 was knocked down by RNA interference, the mRNA expression of CuZn-SOD in the mantle was significantly down-regulated (p < 0.05); a significant positive correlation of expression level between CuZn-SOD and PfAQP4 was found (r = 0.818, p < 0.001). We showed that PfAQP4 plays an important role in the immune response of P. f. martensii, which provide new data for the innate immunity of P. f. martensii.
PAN Xiaolan , LIU Huiru , XU Meng , XU Hanzhi , ZHANG Hua , HE Maoxian . Preliminary study on immunity function of aquaporin AQP4 in the pearl oyster Pinctada fucata martensii[J]. Journal of Tropical Oceanography, 2021 , 40(2) : 83 -89 . DOI: 10.11978/2020050
表1 本研究所用的引物序列Tab. 1 Primers used in this study |
| 引物名称 | 引物序列(5' — 3') | 用途 |
|---|---|---|
| dsAQP4-T7F | GGATCCTAATACGACTCACTATAGGTTGCCTTTGGGCTATCGG | PfAQP4 RNAi含T7启动子序列正向引物 |
| dsAQP4-T7R | GGATCCTAATACGACTCACTATAGGCGGAAGGAGCAGCAAAGAC | PfAQP4 RNAi含T7启动子序列反向引物 |
| dsGFP4-T7F | GGATCCTAATACGACTCACTATAGGCGACGTAAACGGCCACAAGTT | GFP RNAi含T7启动子序列正向引物 |
| dsGFP-T7R | GGATCCTAATACGACTCACTATAGGATGGGGGTGTTCTGCTGGTAG | GFP RNAi含T7启动子序列反向引物 |
| dsAQP4-F | TTGCCTTTGGGCTATCGG | PfAQP4 RNAi正向引物 |
| dsAQP4-R | CGGAAGGAGCAGCAAAGAC | PfAQP4 RNAi反向引物 |
| dsGFP-F | CGACGTAAACGGCCACAAGTT | GFP RNAi正向引物 |
| dsGFP-R | ATGGGGGTGTTCTGCTGGTAG | GFP RNAi反向引物 |
| Faqp | CTGTGATGGCGTCAACTGATG | PfAQP4 qPCR正向引物 |
| Raqp | CGCAATCGTGAGAAGATGACC | PfAQP4 qPCR反向引物 |
| F18S | CGTTTCAACAAGACGCCAGTAG | 18S qPCR正向引物 |
| R18S | ACGAAAAAAAGGTTTGAGAGACG | 18S qPCR反向引物 |
| QT5Rel143 | GATGGCAGAGGATGATTCTTCTT | NF-κB qPCR正向引物 |
| QT3Rel332 | TGATGGACCTTCACACTCATACC | NF-κB qPCR反向引物 |
| MMPF | TCTGGCTCATGCGTTTTTCC | MMP qPCR正向引物 |
| MMPR | AGGGCATGTCCAATCTCATGAG | MMP qPCR反向引物 |
| QT5IL17seq460 | CCAGTCCTCGTAATAAATGTGAACC | IL-17 qPCR正向引物 |
| QT3IL17seq636 | CGCTTCCGCTGCTAGATTCTT | IL-17 qPCR反向引物 |
| LAMPF | TGCCTGTCACAATAATAACC | LAMP qPCR正向引物 |
| LAMPR | AGACTCAAAGTAAGACCACCT | LAMP qPCR反向引物 |
| CuZn-SODF | GAAACTGCAAACCCTATA | CuZn-SOD qPCR正向引物 |
| CuZn-SODR | AGACATTCGTCAAACTCA | CuZn-SOD qPCR反向引物 |
注: 下划线表示T7启动子序列, 粗体表示额外的碱基, 可允许聚合酶更有效的结合和启动 |
图1 3种免疫刺激引起PfAQP4 mRNA相对表达量在外套膜(a)和消化腺(b)中的随时间变化情况*表示同一时刻试验组与对照组有显著差异(p<0.05) Fig. 1 Relative mRNA expression of PfAQP4 in the mantle (a) and digestive gland (b) after immune stimulation * indicates that the experimental group and control group are significantly different at the same time (p < 0.05) |
图2 PfAQP4表达抑制后5个免疫相关基因在外套膜中的随时间变化情况a. PfAQP4 mRNA; b. LAMP mRNA; c. CuZn-SOD mRNA; d. MMP mRNA; e. NF-κB mRNA; f. IL-17 mRNA。*表示对照组与AQP组差异显著(p<0.05), **表示p<0.01, ***表示p<0.001 Fig. 2 Relative mRNA expression of 5 immune-related genes in the mantle after inhibition of PfAQP4 expression * Indicates significant difference between the control group and AQP group (p < 0.05), ** indicates p < 0.01, and *** indicates p < 0.001 |
表2 RNA干扰后PfAQP4与免疫相关基因mRNA表达量的相关性分析Tab. 2 Correlation analysis of mRNA expression between PfAQP4 and immune-related genes in the mantle after RNA interference |
| 基因名称 | 相关系数 | p值 |
|---|---|---|
| 基质金属蛋白分解酶 | 0.356 | 0.212 |
| 核因子κB | 0.655* | 0.011 |
| 白细胞介素17 | 0.515 | 0.060 |
| 溶酶体膜相关糖蛋白 | 0.674** | 0.008 |
| 铜锌超氧化物歧化酶 | 0.818*** | 0.000 |
注: *表示p<0.05, **表示p<0.01, ***表示p<0.001 |
| [1] |
高沿, 2016. 凡纳滨对虾水孔蛋白-4基因克隆、表达及其对盐度胁迫的反应研究[D]. 北京: 中国科学院大学(in Chinese).
|
| [2] |
潘肖兰, 刘惠茹, 许濛, 等, 2020. 马氏珠母贝水通道蛋白基因AQP4 cDNA克隆和表达分析[J]. 热带海洋学报, 39(3):66-75.
|
| [3] |
孙乐, 倪云翔, 丁之德, 2016. 宫内环境变化对子代大脑海马发育影响的分子机制[J]. 国际妇产科学杂志, 43(5):547-551, 560.
|
| [4] |
王晶, 李红玲, 赵龙, 等, 2018. 不同疗程高压氧治疗对脑出血大鼠血肿周围AQP4和SOD表达的影响[J]. 中国康复医学杂志, 33(1):29-35.
|
| [5] |
武柠子, 马慧萍, 王宁, 等, 2016. 脑水肿分子机制的研究进展[J]. 解放军医药杂志, 28(6):14-18.
|
| [6] |
袁牧, 王昌留, 王一斐, 等, 2016. 超氧化物歧化酶的研究进展[J]. 中国组织化学与细胞化学杂志, 25(6):550-558.
|
| [7] |
朱丹, 李宏俊, 高祥刚, 等, 2010. 文蛤胞内铜锌超氧化物歧化酶基因的克隆与序列分析[J]. 生物技术通报,(11): 123-128, 133.
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}