
Journal of Tropical Oceanography >
Analysis of the bloom caused by colonial Phaeocystis globosa in Mirs Bay
Copy editor: LIN Qiang
Received date: 2021-11-01
Revised date: 2021-12-18
Online published: 2021-12-23
Supported by
Natural Science Foundation of China(41606176)
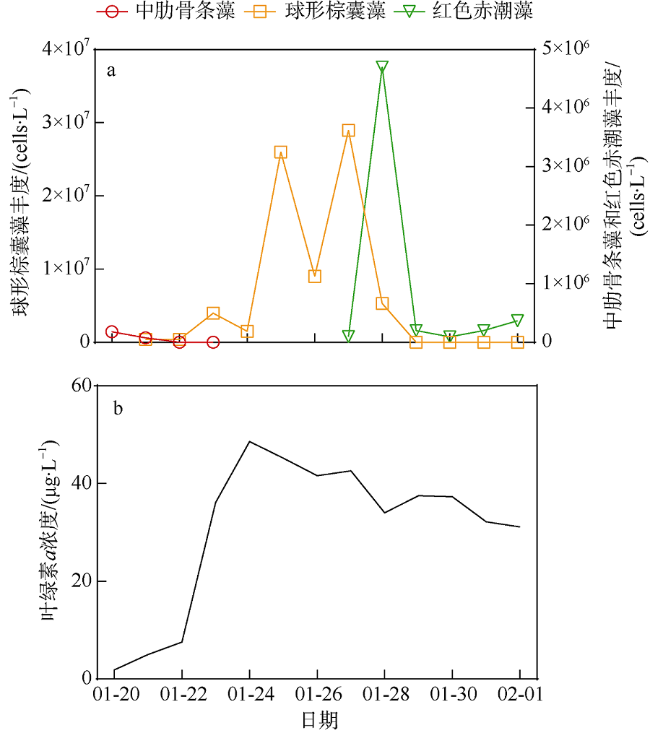
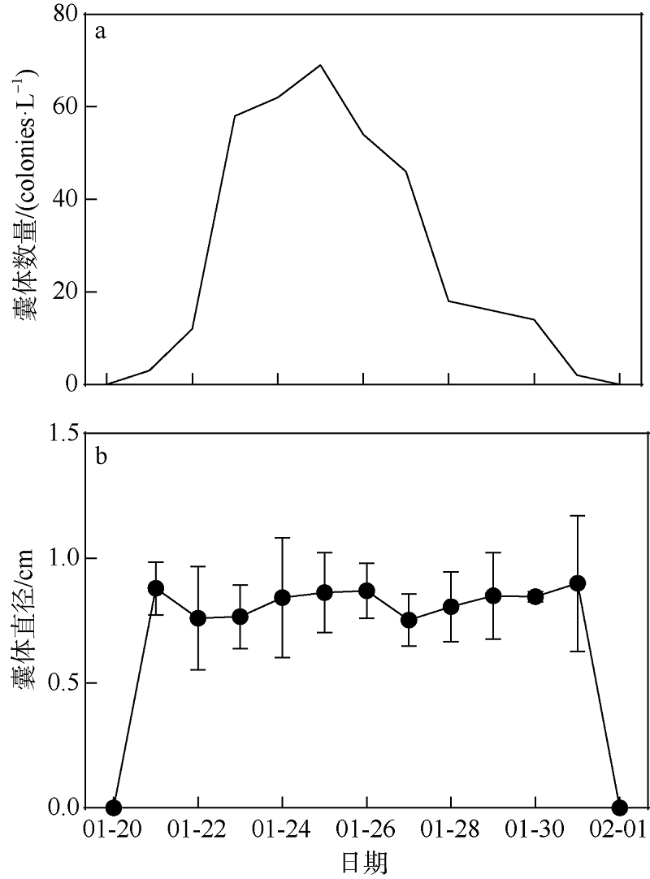
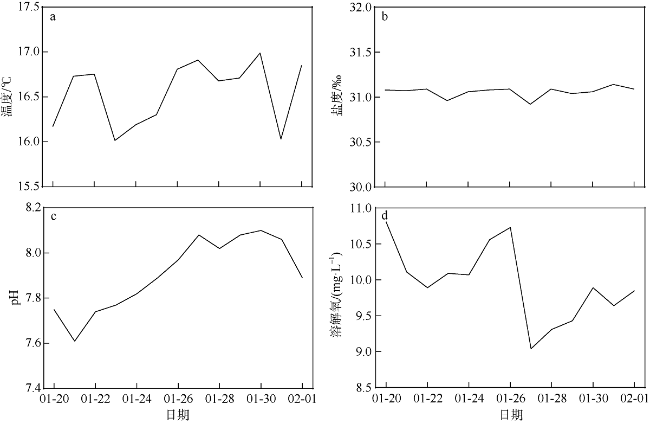
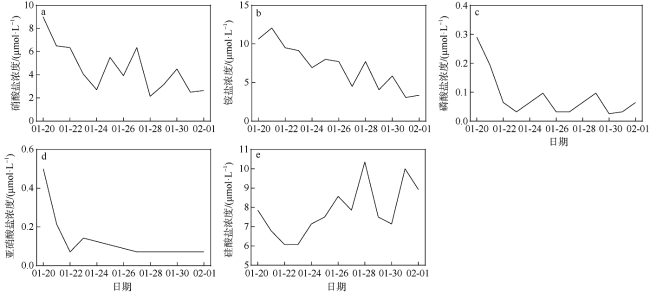
A bloom caused by giant colonies of Phaeocystis globosa was observed in Mirs Bay in late January 2021. To better understand the bloom dynamics and their controlling factors, in situ investigations on phytoplankton succession, colony numbers and size, inorganic nutrient concentrations, and other environmental factors were conducted from Jan. 20 to Feb. 1, 2021. The phytoplankton bloom, initiated by an early Skeletonema costatum bloom, was followed by a large bloom of P. globosa. Phytoplankton community was dominated by the population of Akashiwo sanguinea after the decline of P. globosa bloom. The maximum colony density observed during the P. globosa bloom was up to 69 colonies ·L-1, while colony diameters were up to 1.2 cm. Analytic results showed that ammonium and nitrate were the key environmental factors that initiated the P. globosa bloom. In addition, turbulence and competitive advantage over S. costatum under phosphate limitation might also play important roles in the occurrence of the bloom. The bloom of A. sanguine could be the major cause for the decline of P. globosa bloom. Overall, the P. globosa bloom was a combined result of various physical and chemical factors and biotical factors.
Key words: Mirs Bay; Phaeocystis globosa; bloom; environmental factors
LIU Yue , LI Li , ZHAI Xiaohui , ZHOU Juan , YE Penghao , HUANG Shengdong . Analysis of the bloom caused by colonial Phaeocystis globosa in Mirs Bay[J]. Journal of Tropical Oceanography, 2022 , 41(3) : 164 -171 . DOI: 10.11978/2021148
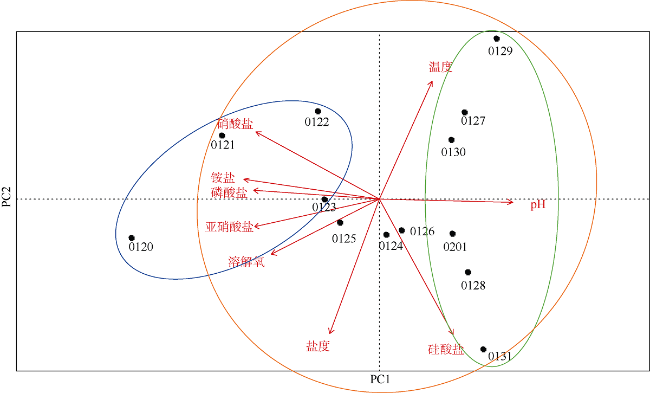
图5 调查期间环境因子主成分分析蓝色表示硅藻藻华期, 橙色表示球形棕囊藻藻华期, 绿色表示甲藻藻华期 Fig. 5 Principal component analysis of environmental parameters in Mirs Bay during the bloom. Blue ellipse represents diatom bloom, orange ellipse represents Phaeocystis globosa bloom, and green ellipse represents dinoflagellate bloom |
表1 球形棕囊藻囊体丰度与环境因子关联度Tab. 1 The value of grey association degree between Phaeocystis globosa colony density and each environmental factor |
| 序号 | 环境因子 | 关联系数 |
|---|---|---|
| 1 | 铵盐 | 0.7024 |
| 2 | 硝酸盐 | 0.6957 |
| 3 | 溶解氧 | 0.6886 |
| 4 | 盐度 | 0.6854 |
| 5 | pH | 0.6847 |
| 6 | 温度 | 0.6836 |
| 7 | 硅酸盐 | 0.6785 |
| 8 | 磷酸盐 | 0.6537 |
| 9 | 亚硝酸盐 | 0.6520 |
| [1] |
蔡卓平, 段璐洋, 肖群, 等, 2011. 风浪扰动促进中肋骨条藻和球形棕囊藻增殖的模拟研究[J]. 海洋环境科学, 30(4): 473-476.
|
| [2] |
陈菊芳, 徐宁, 江天久, 等, 1999. 中国赤潮新记录种──球形棕囊藻(Phaeocystis globosa)[J]. 暨南大学学报(自然科学与医学版), 20(3): 124-129.
|
| [3] |
贺成, 宋书群, 李才文, 2019. 广西北部湾海域球形棕囊藻囊体时空分布及其影响因素[J]. 海洋与湖沼, 50(3): 630-643.
|
| [4] |
黄博珠, 2016. 混合营养型甲藻红色赤潮藻对共存浮游植物的毒性效应[D]. 广州: 暨南大学: 1-59.
|
| [5] |
黄肖阳, 龙寒, 莫钰, 等, 2021. 不同形态磷源对球形棕囊藻生长及囊体形成的影响[J]. 海洋湖沼通报, (1): 116-122.
|
| [6] |
李冬梅, 高永利, 田甜, 等, 2010. 水体扰动对多种赤潮藻生长的影响[J]. 热带海洋学报, 29(6): 65-70.
|
| [7] |
吕旭宁, 2020. 硝酸盐在北部湾球形棕囊藻藻华过程中的关键影响作用[D]. 青岛: 中国科学院大学(中国科学院海洋研究所): 1-137.
|
| [8] |
沈萍萍, 齐雨藻, 欧林坚, 2018. 中国沿海球形棕囊藻(Phaeocystis globosa)的分类、分布及其藻华[J]. 海洋科学, 42(10): 146-162.
|
| [9] |
田晶晶, 2010. 环境因子对球形棕囊藻细胞群体形成的影响[D]. 广州: 暨南大学: 1-69.
|
| [10] |
王超, 李新辉, 赖子尼, 等, 2010. 珠江口球形棕囊藻(Phaeocystis globosa)赤潮后期的浮游植物群落结构特征研究[J]. 生态科学, 29(2): 140-146.
|
| [11] |
王艳, 唐海溶, 2006. 不同形态的磷源对球形棕囊藻生长及碱性磷酸酶的影响[J]. 生态科学, 25(1): 38-40.
|
| [12] |
王艳, 邓坤, 王小冬, 2013. 球形棕囊藻囊体形成中光照、营养盐和共存硅藻的影响[J]. 生态科学, 32(2): 165-170.
|
| [13] |
徐宁, 齐雨藻, 陈菊芳, 等, 2003. 球形棕囊藻(Phaeocystis globosa Scherffel)赤潮成因分析[J]. 环境科学学报, 23(1): 113-118.
|
| [14] |
赵越, 于仁成, 张清春, 等, 2019. 北部湾海域微型、微微型浮游植物类群季节变化及其与棕囊藻赤潮的关系初探[J]. 海洋与湖沼, 50(3): 590-600.
|
| [15] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会 2008. GB 17378.4-2007 海洋监测规范第4部分: 海水分析[S]. 北京: 中国标准出版社.
General Administration of Quality Supervision Inspection and Quarantine of the People's Republic of China, Standardization Administration, 2008. GB 17378.4-2007 The specification for marine monitoring-Part 4: seawater analysis[S]. Beijing: Standards Press of China. (in Chinese)
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}