
Journal of Tropical Oceanography >
Ovarian development, histology of oogenesis and yolk formation of Tapes conspersus from the Beihai Yingpan, Guangxi
Copy editor: YIN Bo
Received date: 2021-12-29
Revised date: 2022-02-16
Online published: 2022-02-18
Supported by
Guangxi Innovation Driven Development Project(AA19254032)
National Basic Research and Development Program of China(2018YFD0901400)
The ovary development, oogenesis histology and ultrastructure of vitellogenesis of Tapes conspersus from the Beihai Yingpan, Guangxi were studied using tissue sectioning and transmission electron microscopy based on the samples taken from November 2020 to October 2021. The results showed that the ovary development of T. conspersus had a one-year cycle, consisting of the proliferating stage (from the end of June to July), the growth stage (from mid-July to mid-October), the maturing stage (from November to the end of February of the following year), and the spawning stage (from March to April) and the resting stage (from May to June). Ovary development is strongly affected by water temperature and seasons. Gonadal development among individuals is not completely synchronized. Gonads begin to develop in the proliferating stage each year, and a large number of mature eggs are discharged at the spawning stage. According to the characteristics of the oocyte cell development of T. conspersus, it can be divided into four stages: oogonium, primary oocyte, secondary oocytes and mature oocyte. Based on the observation of the occurrence of vitellogenesis of T. conspersus by transmission electron microscope, it can be divided into three periods: early period of vitellogenesis, period of vitellogenesis, and late period of vitellogenesis. The results of this study provide basic data for the reproduction biology and artificial breeding of the T. conspersus.
LIAN Changpeng , WU Tao , WANG Chaoqi , YANG Ling , PAN Ying . Ovarian development, histology of oogenesis and yolk formation of Tapes conspersus from the Beihai Yingpan, Guangxi[J]. Journal of Tropical Oceanography, 2022 , 41(5) : 170 -179 . DOI: 10.11978/2021185
表1 广西北海营盘海域钝缀锦蛤卵巢发育各期的周年分布Tab. 1 Annual distribution of ovarian development stages of Tapes conspersus in the Beihai Yingpan, Guangxi |
| 取样日期 | 雌性发育分期的个体数(占比) | 雌性总数/只 | 盐度‰ | 水温/℃ | ||||
|---|---|---|---|---|---|---|---|---|
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | ||||
| 2020-11-29 | 4 (28.6%) | 10 (71.4%) | 14 | 27.4 | 22.0 | |||
| 2020-12-21 | 5 (26.3%) | 14 (73.7%) | 19 | 27.5 | 17.2 | |||
| 2021-01-08 | 2 (16.7%) | 10 (83.3%) | 12 | 29.4 | 15.6 | |||
| 2021-02-24 | 1 (7.1%) | 13 (92.9%) | 14 | 30.9 | 17.3 | |||
| 2021-03-18 | 6 (100%) | 6 | 30.0 | 21.0 | ||||
| 2021-04-22 | 9 (100%) | 9 | 30.0 | 27.5 | ||||
| 2021-05-20 | 2 (25%) | 6 (75%) | 8 | 28.0 | 29.5 | |||
| 2021-06-23 | 1 (12.5%) | 7 (87.5%) | 8 | 28.4 | 30.5 | |||
| 2021-07-15 | 4 (25%) | 12 (75%) | 16 | 29.0 | 29.7 | |||
| 2021-08-18 | 6 (75%) | 2 (25%) | 8 | 30.0 | 29.8 | |||
| 2021-09-17 | 1 (7.1%) | 12 (85.8%) | 1 (7.1%) | 14 | 31.0 | 29.8 | ||
| 2021-10-20 | 7 (70%) | 3 (30%) | 10 | 29.0 | 25.5 | |||
图2 雌性钝缀锦蛤各期生殖细胞组织切片a. 卵原细胞期, 示卵原细胞(Og); b. 初级卵母细胞期, 示初级卵母细胞(Po)、滤泡(F); c. 次级卵母细胞期, 示次级卵母细胞(So)、核仁(Nu); d. 成熟卵母细胞期, 示成熟卵母细胞(Ov)、嗜酸性颗粒(A) Fig. 2 Germ cell tissue sections of female Tapes conspersus at each stage. (a) oogonium period, shows oogonium (Og); (b) primary oocyte period, shows primary oocyte (Po), Follicle (F); (c) secondary oocyte period, shows Secondary oocyte (So), Nucleolus (Nu); (d) mature oocyte, shows Ovum (Ov), Acidophilic granule (A) |
表2 钝缀锦蛤各期雌性生殖细胞特征Tab. 2 The female reproductive cell of Tapes conspersus |
| 分期 | 细胞特点 | 出现时期及位置 | 细胞核特点 |
|---|---|---|---|
| 卵原细胞期 | 呈圆形, 细胞直径约为4.34~6.71µm, 细胞体积小, 在滤泡中排列疏松 | 除休止期外, 其他期均可见, 增殖期和生长期数量最多, 常常附在滤泡壁的基膜上 | 细胞整体染色较深, 可以观察到清晰的细胞核, 核仁在这一时期不明显, 细胞核的位置常位于中央 |
| 初级卵母细胞期 | 常被周围的结缔组织包裹着, 细胞形状多呈圆形, 直径约为9.47~ 10.22µm, 细胞体积较前期的卵原细胞大 | 增殖期及生长期数量最多, 常附于滤泡壁的基膜上 | 可见聚集成团的核仁, 染色质数量少且不清晰, 在细胞中呈絮状。细胞核染色较细胞质浅 |
| 次级卵母细胞期 | 可以清晰观察到此期的卵母细胞卵柄依旧存在, 形状未呈圆形, 常呈梨形, 细胞直径约为18.02~20.73µm | 生长期、成熟期、排放期可见大量此期细胞, 由初级卵母细胞发育成熟而得, 附着于基膜 | 核膜清晰, 染色质逐渐清晰, 数量增多, 互相缠绕, 核仁染色最深, 其次是细胞质, 核仁位于细胞核一侧 |
| 成熟卵母细胞期 | 成熟的卵母细胞呈圆形, 未成熟的呈梨形或者椭圆形等, 细胞直径约为39.57~62.71µm, 经过不断发育, 卵母细胞内的细胞质中充满了大小不一的卵黄颗粒, 此时的卵母细胞的体积达到了最大, 处于成熟待排放阶段 | 滤泡腔中存在大量圆形或者椭圆形、梨形的卵母细胞, 此时卵柄已经脱落。在成熟期、排放期以成熟的卵母细胞为主, 因为大量成熟的卵母细胞挤压在狭小的滤泡腔中, 导致部分卵母细胞变形 | 细胞核体积达到最大直径为23.49~ 25.87µm, 核仁直径约为4.49~ 5.74µm。常有1~2个核仁分布在细胞核中, 细胞整体染色较深 |
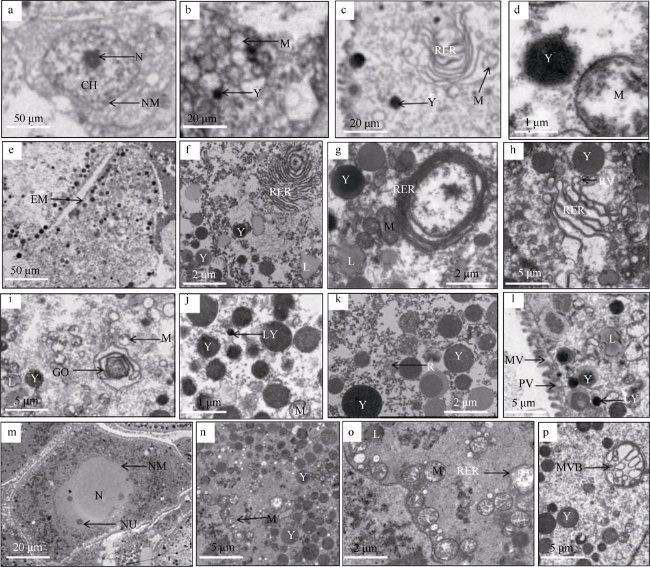
图3 卵母细胞发育的超微结构观察a. 卵黄合成前期卵母细胞, 示细胞核(N)、染色质(CH)、核膜(NM), ×300; b. 卵黄合成前期, 示线粒体(M)、卵黄(Y), ×8000; c. 卵黄合成前期, 示粗面内质网(RER)、线粒体(M)、卵黄(Y), ×8000; d. 卵黄合成前期, 示卵黄(Y)、线粒体(M), ×8000; e. 卵黄合成期卵母细胞, 示细胞膜(EM), ×300; f. 卵黄合成期, 示粗面内质网包绕脂滴形成卵黄粒, 脂滴(L)、卵黄(Y)、粗面内质网(RER), ×2500; g. 卵黄合成期, 示线粒体形成的卵黄粒(MY), 粗面内质网(RER)、脂滴(L)、卵黄(Y)、线粒体(M), ×5000; h. 卵黄合成期, 示片层状粗面内质网两端膨化, 卵黄(Y)、粗面内质网(RER)、粗面内质网囊泡(RV), ×2000; i. 卵黄合成期, 示高尔基复合体液泡形成卵黄粒, 高尔基复合体(GO)、线粒体(M)、脂滴(L)、卵黄(Y), ×2000; j. 卵黄合成期, 示溶酶体(LY)、卵黄(Y)、线粒体(M), ×5000; k. 卵黄合成期, 示核糖体(R)、卵黄(Y), ×5000; l. 卵黄合成期, 示吞饮泡(PV)、微绒毛(MV)、溶酶体(LY)、脂滴(L)、卵黄(Y), ×2000; m. 卵黄合成后期卵母细胞, 示细胞核(N)、核膜(NM)、核仁(NU), ×500; n. 卵黄合成后期, 示线粒体(M)、卵黄(Y), ×2000; o. 卵黄合成后期, 示粗面内质网空泡化, 粗面内质网(RER)、线粒体(M)、脂滴(L), ×5000; p. 卵黄合成后期, 示多泡小体(MVB)、卵黄(Y), ×5000 Fig. 3 Ultrastructure observation of oocyte development. (a) The early period of vitellogenesis oocyte, showing nucleus (N), chromatin (CH), nuclear membrane (NM), ×300; (b) At previtollogenic stage, showing mitochondria (M) and yolk granules (Y), ×8000; (c) At previtollogenic stage, showing rough endoplasmic reticulum (RER), mitochondria (M) and yolk granules (Y), ×8000; (d) At previtollogenic stage, shows yolk granules (Y) and mitochondria (M), ×8000; (e) The period of vitellogenesis oocyte showing cell membrane (EM), ×300; (f) At vitellogenic stage, showing Rough endoplasmic reticulum surrounds lipid droplets to form yolk granules, lipid droplet (L), yolk granules (Y) and rough endoplasmic reticulum (RER), ×2500; (g) At vitellogenic stage, yolk granule formed by mitochondria (MY), rough endoplasmic reticulum (RER), lipid droplet (L), yolk granules (Y) and mitochondria (M), ×5000; (h) At vitellogenic stage, showing that the two ends of the lamellar rough endoplasmic reticulum are expanded, yolk granules (Y), rough endoplasmic reticulum (RER) and rough endoplasmic reticulum vesicles (RV), ×2000; (i) At vitellogenic stage showing golgi complex vacuoles form yolk granules, golgi complex (GO), mitochondria (M), lipid droplet (L) and yolk granules (Y), ×2000; (j) At vitellogenic stage showing lysosome (LY), yolk granules (Y) and mitochondria (M), ×5000; (k) At vitellogenic stage, showing ribosome (R) and yolk granules (Y), ×5000; (l) At vitellogenic stage, showing pinocytosis vesicles (PV), microvilli (MV), lysosome (LY), lipid droplet (L), and yolk granules (Y), ×2000; (m) The late period of vitellogenesis oocyte, shows nucleus (N), nuclear membrane (NM) and nucleolus (NU), ×500; (n) At postvitellogenic stage, shows mitochondria (M) and yolk granules (Y), ×2000; (o) At postvitellogenic stage, showing cavitation of rough endoplasmic reticulum, rough endoplasmic reticulum (RER), mitochondria (M) and lipid droplet (L), ×5000; (p) At postvitellogenic stage, showing multivesicular bodies (MVB), yolk granules (Y), ×5000 |
表3 钝缀锦蛤各期卵母细胞特征Tab. 3 The characteristics of oocytes at different stages of Tapes conspersus |
| 分期 | 细胞特点 | 细胞器特点 | 卵黄特点 |
|---|---|---|---|
| 卵黄合成前期 卵母细胞 | 用卵柄附着在滤泡壁的基膜上, 形态不规则, 细胞体积小, 有较多的数量。核位于中央。染色质呈颗粒状分布在细胞质中 | 细胞质中可以观察到少量线粒体、粗面内质网等细胞器, 线粒体多呈球形或者椭圆形, 部分线粒体内的管状脊少, 有不发达的基质。粗面内质网并不发达, 周围有少量囊泡 | 可观察到少量且体积较小的卵黄颗粒, 此时的卵黄颗粒无膜结构, 内部有一定数量的电子。周围的细胞质中有数量众多的糖原颗粒分布 |
| 卵黄合成期 卵母细胞 | 此期细胞形状不规则, 细胞质内部有大量的细胞器、脂滴等物质, 细胞膜附近积累部分卵黄 | 此期细胞器发达且数量众多。线粒体脊明显, 少量已经转化为卵黄粒。内质网有环状和多层片状两种形式。部分高尔基体在内部会包裹其他物质形成囊泡 | 卵黄颗粒在这一时期大量开始生成, 大多数呈圆球形, 部分形态不规则, 卵黄颗粒有电子密度高的, 也有电子密度低的 |
| 卵黄合成后期 卵母细胞 | 细胞体积达到最大, 细胞核和核仁清晰可见。此期的卵母细胞基本处于成熟阶段, 因相互挤压, 形状不规则, 部分卵母细胞可以观察到2个核仁, 但大多是1个核仁 | 线粒体数量显著增加, 大量分布在细胞质中, 可见部分线粒体内部脊开始消失, 呈现空泡化, 并逐渐演化为卵黄颗粒。内质网、高尔基体等细胞器特征已经不明显, 有部分囊泡存在于细胞质中 | 卵母细胞内可以观察到大量形态各异的卵黄颗粒, 大多数呈球形, 此期的卵黄颗粒数量最多, 体积最大, 密集分布于细胞质中 |
| [1] |
曹伏君, 罗杰, 李长玲, 等, 2010. 细角螺的生殖系统组织学研究[J]. 热带海洋学报, 29(6): 57-64.
|
| [2] |
曹伏君, 刘永, 张春芳, 等, 2012. 施氏獭蛤(Lutraria sieboldii)性腺发育和生殖周期的研究[J]. 海洋与湖沼, 43(5): 976-982.
|
| [3] |
邓传敏, 孔令锋, 于瑞海, 等, 2017. 长牡蛎壳金选育群体性腺发育与营养成分的周年变化[J]. 中国水产科学, 24(1): 40-49.
|
| [4] |
顾海龙, 林志华, 沈伟良, 等, 2013. 泥蚶初级卵母细胞发育及卵黄发生的超微结构研究[J]. 海洋科学, 37(1): 49-53.
|
| [5] |
姜永华, 颜素芬, 2009. 中国龙虾卵子发生及卵黄发生的超微结构观察[J]. 中国水产科学, 16(5): 697-704.
|
| [6] |
刘相全, 方建光, 包振民, 等, 2007. 中国蛤蜊繁殖生物学的初步研究[J]. 中国海洋大学学报, 37(1): 89-92.
|
| [7] |
鹿瑶, 刘辉, 聂鸿涛, 等, 2015. 辽宁沿海薄片镜蛤的繁殖周期研究[J]. 大连海洋大学学报, 30(6): 647-652.
|
| [8] |
吕敏, 李琪, 2022. 山东田横岛海域长牡蛎性腺发育及生化成分周年变化[J]. 中国海洋大学学报(自然科学版), 52(2): 33-40.
|
| [9] |
聂振平, 彭慧婧, 邹杰, 等, 2020. 钝缀锦蛤选育群体F2生长性状相关性及遗传力分析[J]. 广西科学, 27(3): 241-247.
|
| [10] |
宁军号, 常亚青, 宋坚, 等, 2015. 偏顶蛤的性腺发育和生殖周期[J]. 中国水产科学, 22(3): 469-477.
|
| [11] |
区小玲, 苏翔驹, 何俊锋, 等, 2015. 广西北部湾管角螺性腺发育与繁殖规律研究[J]. 中国海洋大学学报, 45(11): 20-28.
|
| [12] |
饶小珍, 陈文列, 陈寅山, 等, 2001. 中国淡水蛏卵子发生的超微结构[J]. 应用与环境生物学报, 7(3): 248-253.
|
| [13] |
王庆恒, 张家炜, 郝瑞娟, 等, 2017. 光裸星虫体腔液中卵子发生的超微结构[J]. 海洋与湖沼, 48(1): 57-66.
|
| [14] |
闻海波, 孙光兴, 丁图强, 等, 2020. 淮河橄榄蛏蚌繁殖类型与性腺发育观察[J]. 中国水产科学, 27(10): 1156-1166.
|
| [15] |
吴明灿, 张立, 潘英, 等, 2015. 糙海参卵子发生及卵黄发生的超微结构[J]. 热带海洋学报, 34(3): 68-74.
|
| [16] |
巫旗生, 文宇, 曾志南, 等, 2017. 钝缀锦蛤繁殖周期和胚胎发育[J]. 中国水产科学, 24(3): 488-496.
|
| [17] |
巫旗生, 曾志南, 宁岳, 等, 2018. 钝缀锦蛤形态性状对活体质量的影响[J]. 水产科学, 37(1): 110-114.
|
| [18] |
杨家林, 邹杰, 彭慧婧, 2019. 温度、盐度和体质量对钝缀锦蛤滤食率和同化率的影响[J]. 水产科学, 38(1): 104-108.
|
| [19] |
应雪萍, 杨万喜, 2001. 泥螺卵黄发生过程中线粒体的变化[J]. 动物学研究, 22(5): 379-382.
|
| [20] |
应雪萍, 2002. 文蛤卵母细胞卵黄发生的超微结构[J]. 中国水产科学, 9(2): 125-128.
|
| [21] |
朱星海, 孙红振, 杨祖晶, 等, 2019. 风信标扇贝的性腺发育与繁殖周期规律研究[J]. 中国海洋大学学报(自然科学版), 49(2): 52-58.
|
| [22] |
庄启谦, 2001. 中国动物志软体动物门双壳纲帘蛤科[M]. 北京: 科学出版社: 63-64. (in Chinese)
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}