
Journal of Tropical Oceanography >
Evolution characteristics and controlling factors of deep-water corals in the southwest Dongsha Sea area since 8000 a BP*
Copy editor: YAO Yantao
Received date: 2022-02-24
Revised date: 2022-04-11
Online published: 2022-04-14
Supported by
The 2020 Research Program of Sanya Yazhou Bay Science and Technology City(SKJC-2020-01-012)
National Natural Science Foundation of China(41976065)
National Natural Science Foundation of China(41776061)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0104)
Deep-water coral (DWC) skeletons record the fine changes in the environment where corals grow. However, the factors influencing the growth of DWCs in the South China Sea, evolutionary stages, and their responses to marine environmental changes are not well understood at present. In this paper, the chemical elements and U-Th ages of DWC skeletons in the Southwest sea area of Dongsha were determined and studied to reveal the development and evolution characteristics of DWCs since 8000 a BP and the factors controlling their growth. Major and trace elements and factor analysis of DWCs show that the growth and evolution of DWCs in the Southwest sea area of Dongsha are influenced by bottom current activity, primary productivity, and terrigenous materials. The evolution of DWCs can be divided into four stages according to U-Th age, Ⅰ: 8000~4500 a BP; Ⅱ: 4500~2500 a BP; Ⅲ: 2500~1200 a BP; Ⅳ: 1200 a BP till now. The drastic fluctuation of bottom current velocity, organic particulate matter, seawater temperature and heat content, and the supply of biogenic materials resulted in the development discontinuity of DWCs at 4500~2500 a BP. The strong bottom current activity, strong winter monsoon, and the supply of organic particulate matter affected the evolution stage and characteristics of DWCs in the southwest sea area of Dongsha. The results provide a new perspective for further exploring the influencing factors, development, and evolution characteristics of DWCs at ocean margins and their responses to ocean processes.
Key words: deep-water coral; U-Th age; evolution stage; Holocene; southwest sea area of Dongsha
ZHANG Bin , CHEN Zhong , XU Antao , WANG Xuesong , TIAN Yuhang , ZHANG Yingwei . Evolution characteristics and controlling factors of deep-water corals in the southwest Dongsha Sea area since 8000 a BP*[J]. Journal of Tropical Oceanography, 2023 , 42(1) : 98 -113 . DOI: 10.11978/2022037
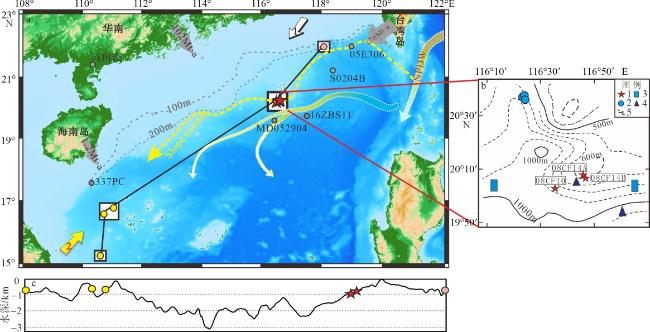
图1 南海北部深水珊瑚分布(a)、研究区地质概况(b)及连接深水珊瑚站位的剖面地形变化(c)该图基于国家测绘地理信息局标准地图服务网站下载的审图号为GS(2021)5447号的标准地图制作。图中五角星为本文珊瑚站位, 黄色圆点为西沙珊瑚站位(Li et al, 2019), 粉色圆点为台西南珊瑚站位(Deng et al, 2019), 灰色圆点为对比研究站位(王雪松 等, 2022; Du et al, 2021; Yang et al, 2020; Wan et al, 2015); 图a中箭头1和2分别为冬季风和夏季风, 灰色箭头及数字代表陆源输入(Liu et al, 2016), 黑色实线为图c剖面路径, NPDW为北太平洋深层水(Wang et al, 2020), slope current为斜坡流(Wang et al, 2020); 图b中图例1代表珊瑚站位, 2代表冷泉碳酸盐岩站位, 3代表泥火山站位, 4代表泥底辟站位, 5代表等深线 Fig. 1 Regional geology and deep water coral stations in the northern South China Sea |
表1 南海北部全新世以来深水珊瑚年龄数据Tab. 1 Age of deep-water corals in the northern South China Sea since the Holocene |
| 样品编号 | 珊瑚类型 | 238U /(×10-9, ±2σ) | 230Th 年龄/(a BP, ±2σ) | 来源 |
|---|---|---|---|---|
| C2 | 石珊瑚 | 4578.4±6.2 | 1990±18 | 本文 |
| C4-1 | 石珊瑚 | 4409.4 ±8.6 | 51±8 | 本文 |
| C4-2 | 石珊瑚 | 4340.0±9.1 | 125±10 | 本文 |
| C5 | 石珊瑚 | 4230. ±8.9 | 47±9 | 本文 |
| C7 | 石珊瑚 | 4330.4±6.7 | 440±9 | 本文 |
| C8-1 | 石珊瑚 | 4164.2±7.9 | 476±9 | 本文 |
| C8-2 | 石珊瑚 | 4377.9±8.3 | 471±9 | 本文 |
| C9 | 石珊瑚 | 4621.4 ±8.8 | 412±9 | 本文 |
| C10 | 石珊瑚 | 4114.7±7.4 | 391±10 | 本文 |
| C11 | 石珊瑚 | 4777.7 ±9.2 | 1122±9 | 本文 |
| C12 | 石珊瑚 | 3735.5±7.1 | 2410±19 | 本文 |
| C13 | 石珊瑚 | 3974.7 ±7.9 | 2435±23 | 本文 |
| C14 | 石珊瑚 | 4555.4±9.2 | 370±9 | 本文 |
| C16 | 石珊瑚 | 4207.6±6.4 | 180±11 | 本文 |
| C17 | 石珊瑚 | 4270.6 ±7.1 | 4664±20 | 本文 |
| C18 | 石珊瑚 | 4553.9±8.2 | 7610±26 | 本文 |
| C19 | 石珊瑚 | 3873.0±6.3 | 2476±22 | 本文 |
| C35 | 石珊瑚 | 4885.6±13.5 | 96±25 | 本文 |
| C36 | 石珊瑚 | 4675.2±8.2 | 55±23 | 本文 |
| C37 | 石珊瑚 | 4461.2±10.4 | 749±43 | 本文 |
| C38 | 石珊瑚 | 4552.4±8.5 | 437±26 | 本文 |
| SY067-9-3 | 柳珊瑚 | 25.6±0.1 | 1 010±237 | 黄恩清 等, 2019 |
| SY068-7-2 | 柳珊瑚 | 47.8±0.1 | 9387±3148 | 黄恩清 等, 2019 |
| SY075-16-2 | 石珊瑚 | 3799.7±8.5 | 718.0±44 | 黄恩清 等, 2019 |
| SY076-9 | 石珊瑚 | 3830.7±5.5 | 6154±94 | 黄恩清 等, 2019 |
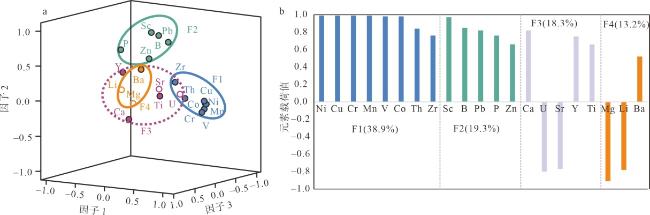
表2 深水珊瑚的常量和微量元素含量Tab. 2 Major and trace elements contents of deep-water corals in the northern South China Sea |
| 序号 | Ca | Mg | Mn | P | Ti | Li | B | Sc | V | Cr | Co | Ni | Cu | Zn | Sr | Y | Zr | Ba | Pb | Th | U |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C2 | 0.39 | 0.60 | 0.45 | 82.39 | 0.41 | 0.69 | 37.55 | 0.60 | 0.10 | 0.82 | 0.96 | 11.42 | 0.27 | 3.04 | 8.46 | 0.22 | 0.01 | 16.75 | 0.41 | 0.001 | 4.37 |
| C4-1 | 0.39 | 0.56 | 1.13 | 76.48 | 0.39 | 0.69 | 36.04 | 0.59 | 0.11 | 0.79 | 0.98 | 11.12 | 0.27 | 2.96 | 8.64 | 0.23 | 0.01 | 16.77 | 0.35 | 0.001 | 4.38 |
| C4-2 | 0.39 | 0.59 | 1.19 | 77.98 | 0.43 | 0.70 | 37.98 | 0.60 | 0.10 | 0.80 | 0.99 | 11.24 | 0.28 | 5.08 | 8.64 | 0.22 | 0.01 | 17.02 | 0.49 | 0.001 | 4.47 |
| C5 | 0.39 | 0.60 | 2.44 | 89.67 | 0.58 | 0.65 | 29.79 | 0.57 | 0.14 | 0.80 | 1.07 | 11.76 | 0.27 | 3.38 | 8.45 | 0.26 | 0.02 | 18.62 | 0.53 | 0.002 | 4.36 |
| C7 | 0.39 | 0.62 | 0.96 | 133.1 | 0.63 | 0.69 | 35.21 | 0.66 | 0.11 | 1.05 | 1.09 | 11.83 | 0.42 | 4.35 | 8.44 | 0.31 | 0.02 | 19.51 | 0.53 | 0.002 | 4.24 |
| C8-1 | 0.38 | 0.62 | 0.97 | 98.35 | 0.49 | 0.71 | 31.81 | 0.61 | 0.12 | 0.77 | 1.01 | 11.38 | 0.35 | 2.98 | 8.56 | 0.27 | 0.02 | 16.82 | 0.47 | 0.001 | 4.04 |
| C8-2 | 0.38 | 0.59 | 0.58 | 70.49 | 0.38 | 0.68 | 25.33 | 0.49 | 0.09 | 0.62 | 1.00 | 11.71 | 0.24 | 2.45 | 8.82 | 0.24 | 0.01 | 17.28 | 0.35 | 0.001 | 4.48 |
| C9 | 0.39 | 0.67 | 0.72 | 94.21 | 0.44 | 0.75 | 31.94 | 0.61 | 0.12 | 0.82 | 1.00 | 11.68 | 0.36 | 3.08 | 8.26 | 0.22 | 0.01 | 16.29 | 0.64 | 0.001 | 3.49 |
| C10 | 0.39 | 0.58 | 0.73 | 73.19 | 0.38 | 0.68 | 28.07 | 0.52 | 0.13 | 0.67 | 1.01 | 12.47 | 0.32 | 3.16 | 8.62 | 0.23 | 0.01 | 16.99 | 0.43 | 0.001 | 4.27 |
| C11 | 0.39 | 0.60 | 0.49 | 82.68 | 0.42 | 0.68 | 35.24 | 0.63 | 0.10 | 0.85 | 1.01 | 11.60 | 0.30 | 3.48 | 8.39 | 0.23 | 0.01 | 17.24 | 0.45 | 0.001 | 4.12 |
| C12 | 0.38 | 0.56 | 0.53 | 91.14 | 0.45 | 0.65 | 32.58 | 0.64 | 0.10 | 0.83 | 1.01 | 11.69 | 0.36 | 3.22 | 8.73 | 0.27 | 0.01 | 17.21 | 0.56 | 0.001 | 4.65 |
| C13 | 0.39 | 0.67 | 0.79 | 129.9 | 0.76 | 0.86 | 39.26 | 0.70 | 0.14 | 1.07 | 1.00 | 11.27 | 0.94 | 3.32 | 8.20 | 0.45 | 0.03 | 17.19 | 0.45 | 0.002 | 3.17 |
| C14 | 0.39 | 0.55 | 0.95 | 114.2 | 0.77 | 0.63 | 31.61 | 0.73 | 0.11 | 0.94 | 1.10 | 12.19 | 0.58 | 5.54 | 8.37 | 0.47 | 0.03 | 17.84 | 0.6 | 0.003 | 3.6 |
| C16 | 0.39 | 0.53 | 2.12 | 132.0 | 0.95 | 0.61 | 31.94 | 0.74 | 0.12 | 0.97 | 1.17 | 12.17 | 0.67 | 7.64 | 8.41 | 0.47 | 0.04 | 20.02 | 0.72 | 0.005 | 3.92 |
| C17 | 0.40 | 0.58 | 23.54 | 69.35 | 1.19 | 0.57 | 16.74 | 0.49 | 0.86 | 381.2 | 2.40 | 145.4 | 14.27 | 4.28 | 8.07 | 0.32 | 0.04 | 14.58 | 0.44 | 0.006 | 3.73 |
| C18 | 0.39 | 0.55 | 35.96 | 98.02 | 0.91 | 0.61 | 34.34 | 0.68 | 1.01 | 464.8 | 3.27 | 229 | 23.08 | 4.81 | 8.39 | 0.32 | 0.06 | 19.88 | 0.61 | 0.01 | 4.32 |
| C19 | 0.38 | 0.57 | 13.95 | 113.4 | 0.76 | 0.63 | 51.15 | 0.90 | 0.37 | 168.6 | 1.87 | 92.59 | 9.08 | 6.68 | 8.51 | 0.33 | 0.03 | 17.19 | 1.18 | 0.003 | 4.43 |
| 最大值 | 0.40 | 0.67 | 35.96 | 133.1 | 1.19 | 0.86 | 51.15 | 0.90 | 1.01 | 464.8 | 3.27 | 229.0 | 23.08 | 7.64 | 8.82 | 0.47 | 0.06 | 20.02 | 1.18 | 0.01 | 4.65 |
| 最小值 | 0.38 | 0.53 | 0.45 | 69.35 | 0.38 | 0.57 | 16.74 | 0.49 | 0.09 | 0.62 | 0.96 | 11.12 | 0.24 | 2.45 | 8.07 | 0.22 | 0.01 | 14.58 | 0.35 | 0.001 | 3.17 |
| 平均值 | 0.39 | 0.59 | 5.15 | 95.68 | 0.61 | 0.68 | 33.33 | 0.63 | 0.23 | 60.38 | 1.29 | 37.09 | 3.06 | 4.09 | 8.47 | 0.30 | 0.02 | 17.48 | 0.54 | 0.002 | 4.12 |
注: Ca含量单位为g·g-1, Mg、Sr含量单位为mg·g-1, 其余元素含量单位为μg·g-1 |
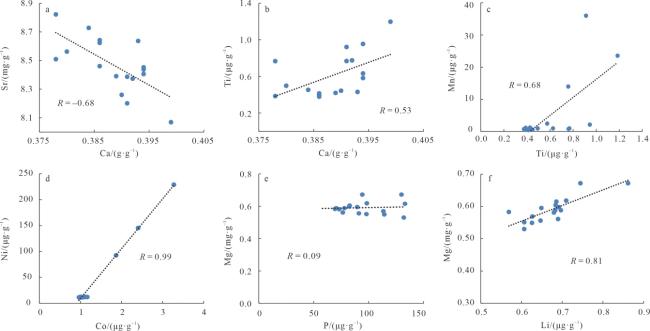
表3 深水珊瑚常量与微量元素间的皮尔逊相关性Tab. 3 Correlation of major and trace elements of deep-water corals in the northern South China Sea |
| 序号 | Ca | Mg | Mn | P | Ti | Li | B | Sc | V | Cr | Co | Ni | Cu | Zn | Sr | Y | Zr | Ba | Pb | Th | U |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ca | 1.00 | ||||||||||||||||||||
| Mg | 0.02 | 1.00 | |||||||||||||||||||
| Mn | 0.21 | -0.30 | 1.00 | ||||||||||||||||||
| P | 0.18 | 0.09 | -0.07 | 1.00 | |||||||||||||||||
| Ti | 0.53* | -0.26 | 0.68** | 0.40 | 1.00 | ||||||||||||||||
| Li | -0.17 | 0.81** | -0.52* | 0.18 | -0.46 | 1.00 | |||||||||||||||
| B | -0.37 | 0.06 | -0.10 | 0.44 | -0.20 | 0.30 | 1.00 | ||||||||||||||
| Sc | -0.13 | -0.18 | 0.13 | 0.75** | 0.32 | -0.05 | 0.78** | 1.00 | |||||||||||||
| V | 0.29 | -0.24 | 0.98** | -0.12 | 0.71** | -0.50* | -0.22 | 0.02 | 1.00 | ||||||||||||
| Cr | 0.24 | -0.27 | 0.99** | -0.12 | 0.70** | -0.52* | -0.18 | 0.06 | 0.99** | 1.00 | |||||||||||
| Co | 0.22 | -0.32 | 0.99** | -0.02 | 0.70** | -0.54* | -0.09 | 0.16 | 0.98** | 0.99** | 1.00 | ||||||||||
| Ni | 0.18 | -0.29 | 0.99** | -0.07 | 0.66** | -0.50* | -0.08 | 0.13 | 0.98** | 0.99** | 0.99** | 1.00 | |||||||||
| Cu | 0.19 | -0.28 | 0.99** | -0.05 | 0.67** | -0.49* | -0.07 | 0.15 | 0.98** | 0.99** | 0.99** | 1.00** | 1.00 | ||||||||
| Zn | 0.27 | -0.52* | 0.30 | 0.55* | 0.62** | -0.50* | 0.32 | 0.71** | 0.23 | 0.26 | 0.33 | 0.28 | 0.29 | 1.00 | |||||||
| Sr | -0.68** | -0.35 | -0.37 | -0.32 | -0.69** | -0.02 | 0.15 | -0.15 | -0.45 | -0.42 | -0.38 | -0.36 | -0.37 | -0.23 | 1.00 | ||||||
| Y | 0.33 | -0.21 | 0.13 | 0.76** | 0.71** | -0.08 | 0.06 | 0.57* | 0.13 | 0.12 | 0.17 | 0.12 | 0.14 | 0.64** | -0.45 | 1.00 | |||||
| Zr | 0.36 | -0.27 | 0.81** | 0.47 | 0.88** | -0.36 | 0.04 | 0.46 | 0.79** | 0.78** | 0.84** | 0.81** | 0.82** | 0.59* | -0.54* | 0.65** | 1.00 | ||||
| Ba | 0.15 | -0.34 | 0.09 | 0.61** | 0.15 | -0.16 | 0.25 | 0.41 | -0.01 | -0.01 | 0.13 | 0.08 | 0.09 | 0.45 | 0.13 | 0.38 | 0.41 | 1.00 | |||
| Pb | -0.18 | -0.22 | 0.28 | 0.49* | 0.36 | -0.30 | 0.59* | 0.84** | 0.18 | 0.23 | 0.31 | 0.28 | 0.29 | 0.72** | -0.12 | 0.34 | 0.42 | 0.24 | 1.00 | ||
| Th | 0.41 | -0.42 | 0.90** | 0.20 | 0.82** | -0.55 | -0.15 | 0.23 | 0.88** | 0.88** | 0.92** | 0.89** | 0.90** | 0.51* | -0.45 | 0.45 | 0.94** | 0.38 | 0.28 | 1.00 | |
| U | -0.43 | -0.52* | 0.03 | -0.42 | -0.43 | -0.40 | 0.16 | -0.12 | -0.05 | -0.01 | 0.02 | 0.04 | 0.02 | -0.07 | 0.78** | -0.58* | -0.30 | 0.18 | 0.03 | -0.12 | 1.00 |
注: *在 0.05 级别, 表示相关性显著; **在 0.01 级别, 表示相关性极显著 |
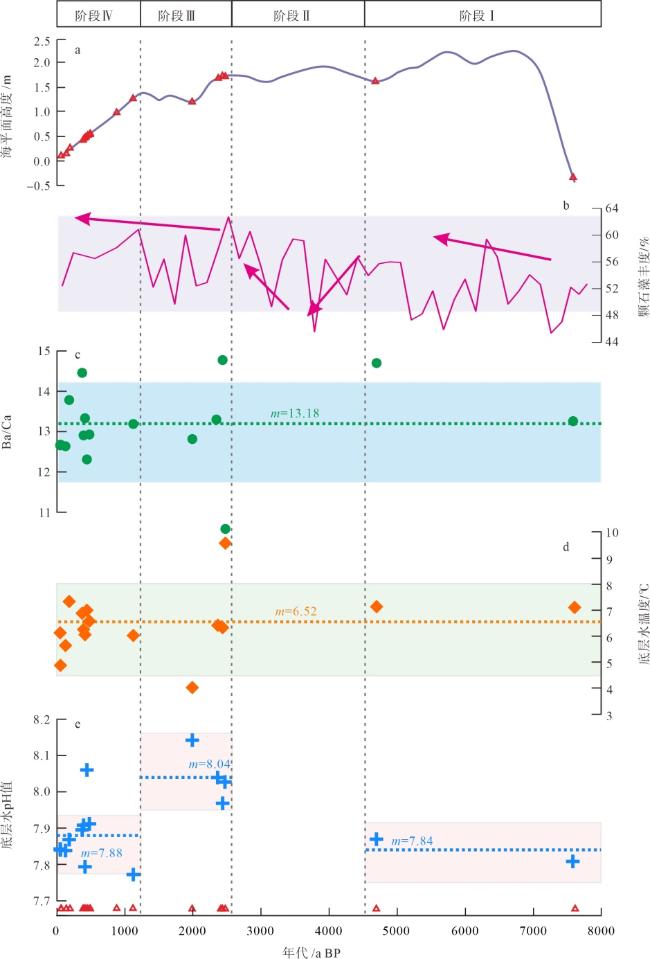
图4 8000a BP以来深水珊瑚的地球化学特征及其重建的环境变化a. 南海北部海平面高度, 据余克服(2012); b. 南海东北部MD052904站位的颗石藻含量变化, 据Su等(2013); c. 本文深水珊瑚骨骼的Ba/Ca(μmol/mol)比值变化, Ba/Ca营养盐指标据Raddatz等(2014); d. 深水珊瑚重建的底层水温度变化, 温度重建公式为Li/Mg (mmol/mol)=5.41exp(-0.049T), 据Montagna等(2014); e. 深水珊瑚重建的底层水pH值变化, pH值重建公式为pH=(U/Ca-16.18)/ (-1.82), 据Raddatz等(2014)。图中红色三角形代表东沙西南海域的深水珊瑚发育年龄; 水平虚线及m表示各阶段参数的平均值 Fig. 4 Geochemical characteristics and reconstructed environmental changes of deep-water corals since 8000 a BP |
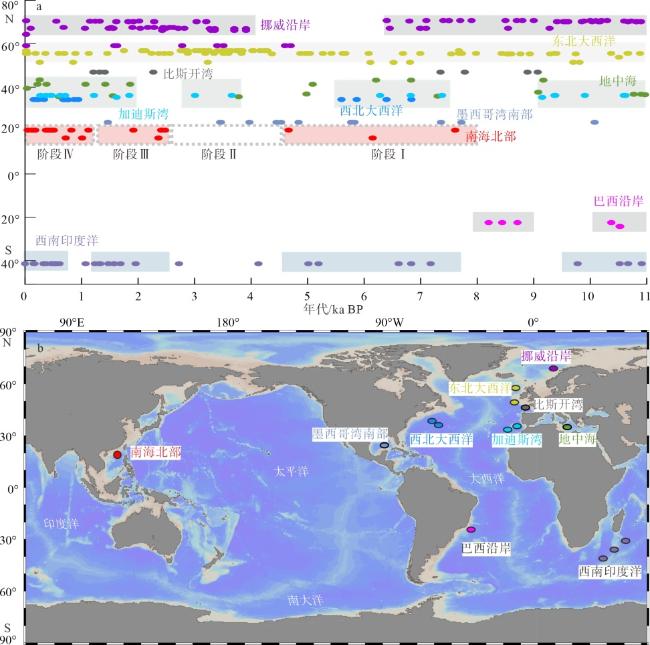
图5 全新世以来南海及其他典型海域深水珊瑚年龄分布与阶段性a. 典型海域的珊瑚生长年龄分布; b. 图a中典型深水珊瑚海域的地理分布。图中东北大西洋数据源自Frank等(2009)和Douarin等(2013); 加迪斯湾数据源自Wienberg等(2010); 巴西沿岸数据源自Mangini等(2010); 挪威沿岸数据源自Wienberg等(2009); 西北大西洋数据源自Matos等(2015); 西南印度洋数据源自Pratt等(2019); 比斯开湾数据源自Eynaud等(2022); 南海北部数据源自本文和黄恩清等(2019) Fig. 5 Age distribution and evolution stages of deep-water corals in the South China Sea and other typical sea areas since Holocene |
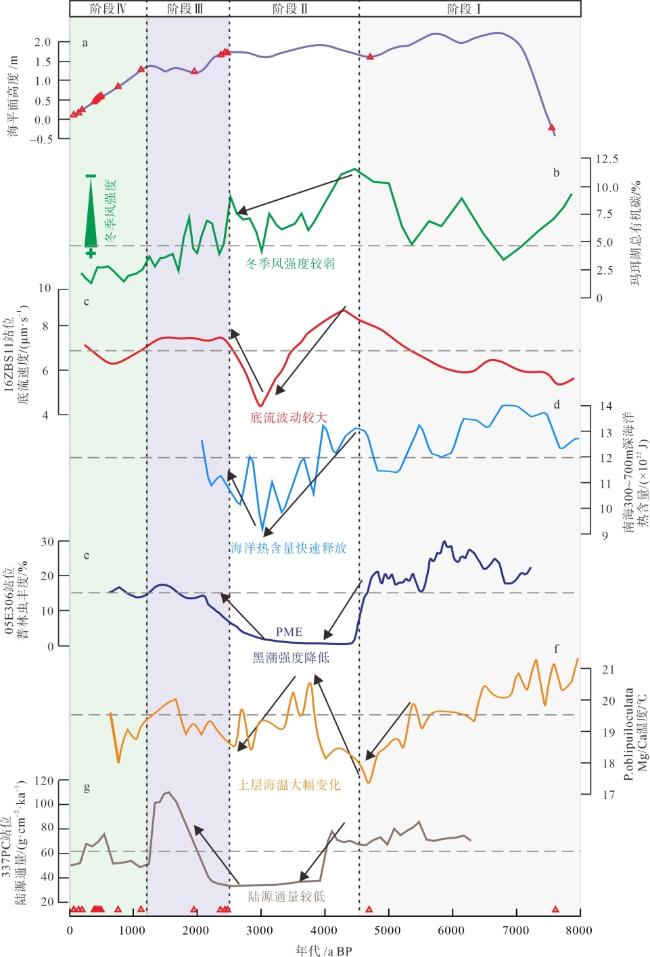
图6 东沙西南海域深水珊瑚演化阶段及影响因素变化图a. 南海北部海平面变化曲线(余克服, 2012); b. 玛珥湖总有机碳变化曲线(Yancheva et al, 2007); c. 南海东北部16ZBS1站位底流速度变化(王雪松 等, 2022); d. 南海水深300~700m处海洋热含量变化曲线(Yang et al, 2020); e. 南海北部05E306站位普林虫丰度变化(Du et al, 2021); f. 南海北部有孔虫Mg/Ca重建的水体温度(Yang et al, 2020); g. 南海北部陆源输入通量变化(Wan et al, 2015); 图中红色三角形代表深水珊瑚发育年龄 Fig. 6 Evolution stages and influencing factors of deep-water corals in the southwest sea area of Dongsha |
| [1] |
安阳, 翦知湣, 2009. 末次冰消期南海南部的普林虫低值事件[J]. 科学通报, 54(17): 2527-2532.
|
| [2] |
蔡观强, 邱燕, 彭学超, 等, 2010. 南海西南海域表层沉积物中微量元素Ba的地球化学特征[J]. 现代地质, 24(3): 560-569.
|
| [3] |
陈建芳, 郑连福,
|
| [4] |
陈越, 王跃, 党皓文, 等, 2021. 南海东北部末次冰盛期以来的水文气候变化[J]. 第四纪研究, 41(4): 1031-1043.
|
| [5] |
陈忠, 杨华平, 黄奇瑜, 等, 2008. 南海东沙西南海域冷泉碳酸盐岩特征及其意义[J]. 现代地质, 22(3): 382-389.
|
| [6] |
陈忠, 莫爱彬, 赵美霞, 等, 2018. 南海北部冷泉区深水珊瑚的发现及其特征[J]. 热带海洋学报, 37(1): 64-71.
|
| [7] |
党皓文, 马小林, 杨策, 等, 2019. 重建高分辨率深海环境变化: 冷水竹节珊瑚无机地球化学方法[J]. 地球科学进展, 34(12): 1262-1272.
|
| [8] |
黄恩清, 孔乐, 田军, 2019. 冷水珊瑚测年与大洋中-深层水碳储库[J]. 地球科学进展, 34(12): 1243-1251.
|
| [9] |
姜大膀, 田芝平, 王娜, 等, 2022. 末次冰盛期和中全新世气候模拟分析进展[J]. 地球科学进展, 37(1): 1-13, doi: 10.11867/j. issn.1001-8166.2021.098.
|
| [10] |
邵磊, 李学杰, 耿建华, 等, 2007. 南海北部深水底流沉积作用[J]. 中国科学 D辑: 地球科学, 37(6): 771-777.
|
| [11] |
王东晓, 肖劲根, 舒业强, 等, 2016. 南海深层环流与经向翻转环流的研究进展[J]. 中国科学: 地球科学, 46(10): 1317-1323.
|
| [12] |
王东晓, 王强, 蔡树群, 等, 2019. 南海中深层动力格局与演变机制研究进展[J]. 中国科学: 地球科学, 49(12): 1919-1932.
|
| [13] |
王雪松, 陈忠, 许安涛, 等, 2022. 南海东北部深海盆末次冰盛期以来陆源碎屑粒度特征及影响因素[J]. 热带海洋学报, 41(1): 158-170.
|
| [14] |
许安涛, 2019. 南海东沙海域冷水珊瑚发育演化的成因模式及其对洋流模态转变的响应[D]. 广州: 中国科学院南海海洋研究所: 1-90.
|
| [15] |
阎贫, 王彦林, 于俊辉, 等, 2021. 东沙海区泥火山调查进展[J]. 热带海洋学报, 40(3): 34-43.
|
| [16] |
余克服, 2012. 南海珊瑚礁及其对全新世环境变化的记录与响应[J]. 中国科学: 地球科学, 42(8): 1160-1172.
|
| [17] |
邹建军, 石学法, 2017. 末次冰期以来北太平洋中层水演化: 研究进展与展望[J]. 地学前缘, 24(4): 141-151.
|
| [18] |
邹仁林, 蒙致民, 关锡廉, 1983. 南海北部大陆架深水石珊瑚的生态分析[J]. 热带海洋, 2(3): 238-243.
ZOU RENLIN, MENG, ZHIMIN,
|
| [19] |
邹仁林, 1988. 南海深水石珊瑚的研究——Ⅱ. 种属记述及时空分布特点[J]. 热带海洋, (1): 76-85.
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}