
Journal of Tropical Oceanography >
Cloning and expression analysis of Cinnamate-4-hydroxylase gene from Rhizophora stylosa
Copy editor: LIN Qiang
Received date: 2021-12-29
Revised date: 2022-03-31
Online published: 2022-04-14
Supported by
National Key Research and Development Program of China(2017FY100700)
National Natural Science Foundation of China(U1901211)
National Natural Science Foundation of China(41876126)
International Partnership Program of Chinese Academy of Sciences(133244KYSB20180012)
Strategic Priority Research Program of the Chinese Academy of Sciences(XDA23050200)
Strategic Priority Research Program of the Chinese Academy of Sciences(XDA19060201)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0305)
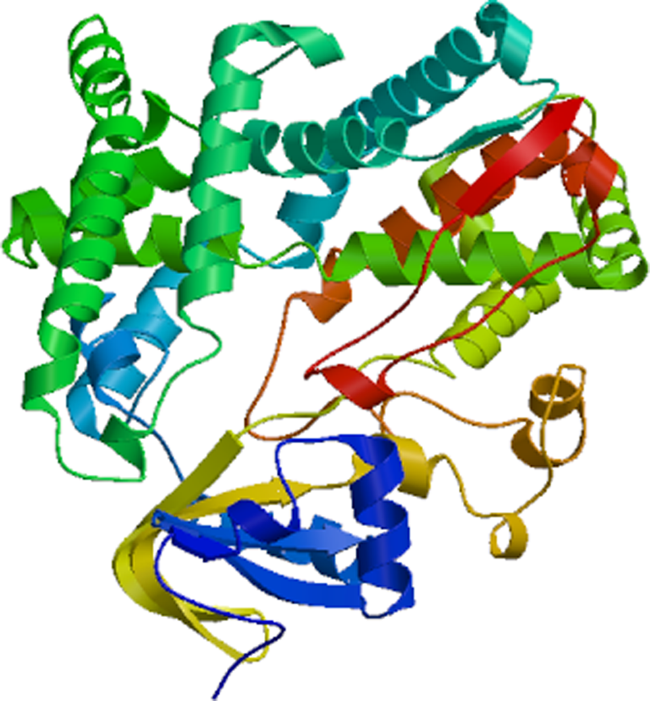
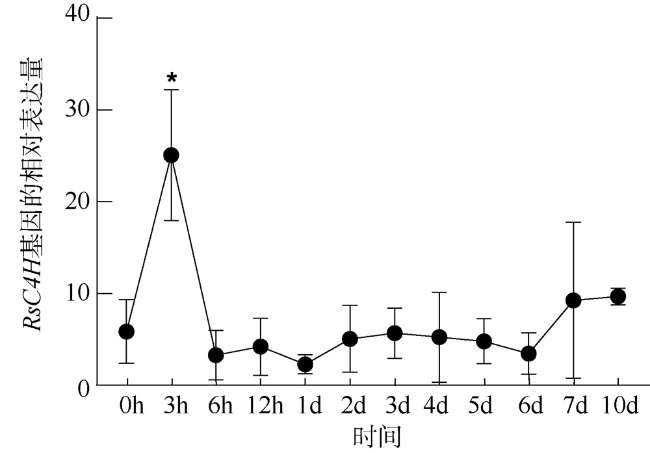
Cinnamate-4-hydroxylase is a key enzyme in lignin synthesis which plays an important role in plant tolerance to heavy metal stress. In this study, a cinnamate-4-hydroxylase gene named RsC4H was cloned from the leaves of Rhizophora stylosa by homologous cloning and rapid amplification of cDNA ends (RACE). Bioinformatics analysis showed that the full length of RsC4H gene cDNA was 2006bp, the length of open reading frame was 1572bp, and encoded 523 amino acids. The relative molecular weight of the encoded protein is 60.18kD. It is a hydrophilic unstable protein with secondary structure alpha-helix (47.42%) and random coil (32.70%), this protein contains a transmembrane structure at the N-terminal and C-terminal, respectively, and does not contain signal peptides. It is mainly distributed on the membrane structure or endoplasmic reticulum to function, and belongs to P450 superfamily. RsC4H protein has high similarity with C4H protein of other plants. The phylogenetic tree results showed that it was closer to Cucurbita maxima (xp_023007159.1) and Cucurbita moschata (xp_022947682.1). qRT-PCR results showed that Rhizophora stylosa could quickly respond to heavy metal stress, improve the expression of RsC4H gene, promote lignin production, increase cell wall thickness and prevent metal ions from entering cells. The results enrich the gene pool of mangrove plants resistant to heavy metal stress, and lay a foundation for revealing the mechanism of heavy metal stress tolerance of mangrove at the molecular level.
XIE Yong , WANG Youshao , ZHANG Weishi . Cloning and expression analysis of Cinnamate-4-hydroxylase gene from Rhizophora stylosa[J]. Journal of Tropical Oceanography, 2022 , 41(6) : 20 -27 . DOI: 10.11978/2021184
表1 引物列表Tab.1 Primer list |
| 引物名称 | 引物序列(5′ to 3′) |
|---|---|
| C4H-F(中间片段) | AAGGGCCAGGACATGGTNTTYAC |
| C4H-R(中间片段) | CTCGTTGATCTCNCCYTTYTGYTG |
| C4H-GSP-5′ | TCACGCTCTTGGTGCTTC |
| C4H-NGSP-5′ | CAAAGCTCTGTGCCAATC |
| C4H-GSP-3′ (RACE) | GCAGTACCGATATGGTTGGG |
| C4H-NGSP-3′ (RACE) | GGGATTGTGCTCAGAAGAAG |
| UPM | CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT |
| UPM short | CTAATACGACTCACTATAGGGC |
| 18S rRNA-F(qRT-PCR) | ATTGGAGGGCAAGTCTGGTG |
| 18S rRNA-R(qRT-PCR) | GATCGACCCATCCCAAGGTC |
| C4H-F(qRT-PCR) | TTGATAGCAGGTTTGAGAGTGAGG |
| C4H-R(qRT-PCR) | CCTCTCAAGAATGGTCTCAAAATG |
表2 生物信息学分析软件或网址Tab. 2 Bioinformatics analysis software or website |
| 预测项目 | 软件版本/网址 |
|---|---|
| 同源比对 | Nucleotide BLAST |
| 开放阅读框查找 | http://www.bio-soft.net/sms/index.html |
| 理化性质预测 | https://web.expasy.org/protparam/ |
| 保守结构域查找 | https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi |
| 信号肽分析 | http://www.cbs.dtu.dk/services/SignalP-4.0/ |
| 跨膜结构查询 | http://www.cbs.dtu.dk/services/TMHMM/ |
| 二级结构 | https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html |
| 三维模型 | https://swissmodel.expasy.org/ |
| 亚细胞定位 | https://wolfpsort.hgc.jp/ |
| 系统发育树构建 | MEGA6 |
| [1] |
黄利娜, 吴光斌, 匡凤元, 等, 2020. 莲雾果实C4H基因的克隆及在NO处理下的表达分析[J]. 集美大学学报(自然科学版), 25(2): 105-112.
|
| [2] |
梁惠桢, 朱家红, 戴好富, 等, 2018. 海南龙血树肉桂酸-4-羟基化酶基因(DcC4H)的克隆及表达分析[J]. 分子植物育种, 16(24): 7984-7989.
|
| [3] |
芮海云, 刘清泉, 沈振国, 2019. 质外体蛋白质在植物重金属耐性中的作用[J]. 生物学杂志, 36(3): 88-91.
|
| [4] |
宋晖, 王友绍, 2012. 萘胁迫下秋茄MnSOD基因和C4H基因的实时定量表达分析[J]. 生态科学, 31(2): 104-108.
|
| [5] |
宋慕波, 周伟政, 唐永胜, 等, 2020. 荸荠肉桂酸4-羟基化酶基因的克隆及其在鲜切荸荠黄化过程中的表达分析[J]. 基因组学与应用生物学, 39(2): 666-673.
|
| [6] |
宋西红, 郝磊, 吕晓玲, 等, 2015. 紫苏肉桂酸4-羟基化酶基因的克隆与表达[J]. 广东农业科学, 42(11): 124-129.
|
| [7] |
王友绍, 孙翠慈, 王玉图, 等, 2019. 生态学理论与技术创新引领我国热带、亚热带海洋生态研究与保护[J]. 中国科学院院刊, 24(1): 121-129.
|
| [8] |
王友绍, 2019. 红树林分子生态学[M]. 北京: 科学出版社: 23-24.
|
| [9] |
姚胜波, 王文钊, 李明卓, 等, 2015. 茶树肉桂酸4-羟基化酶基因的克隆及表达分析[J]. 茶叶科学, 35(1): 35-44.
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}