
Journal of Tropical Oceanography >
Effect of Vibrio harveyi on the quantity and bacterial communities in different tissues of Haliotis discus hannai
Copy editor: YAO Yantao
Received date: 2022-02-18
Revised date: 2022-04-16
Online published: 2022-04-22
Supported by
National Natural Science Foundation of China(31902416)
Research Project of Department of Education of Guangdong Province(2021ZDZX2066)
Huizhou College Research and Innovation Group Cultivation Project(HZU201807)
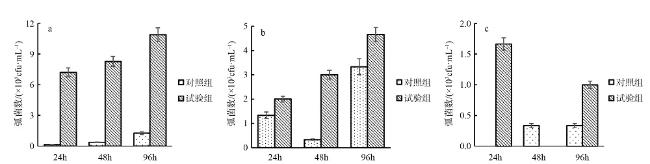
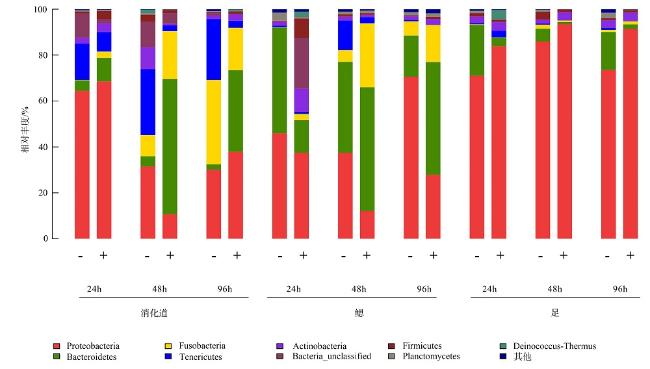
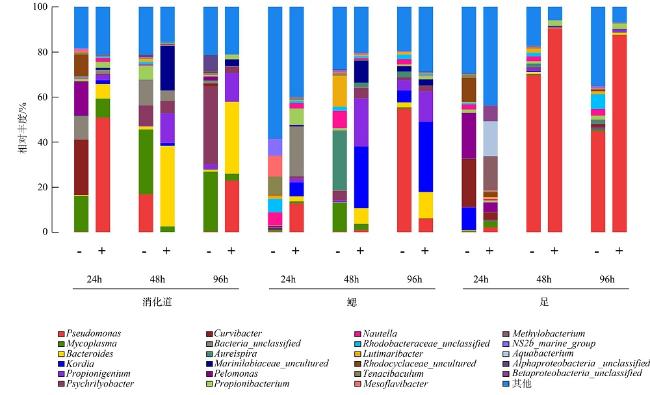
The quantity and the bacterial communities in the digestive tract, gill, and foot of Haliotis discus hannai after infection with Vibrio harveyi were studied using Illumina sequencing technology. Results showed that nine phyla of bacteria were detected in the digestive tract, gills, and foot muscles of H.discus hannai, with Proteobacteria being the dominant phylum, accounting for 30.1%~64.4%, 37.45%~70.6%, and 71.1%~85.9% of the tissues. Bacteroidetes (5.53%~46.09%), Fusobacteria (4.43%~36.80%) and Tenericutes (0.60%~28.77%) were the sub-dominant phyla, while five other phyla including Actinobacteria and some unknown taxa were also detected. At the genus level, the dominant genera in the digestive tract of H.discus hannai were Mycoplasma (15.85%~34.55%) and in the gill was Pseudomonas (54.81%). The dominant genera in the foot muscles were Pseudomonas (44.83%~69.76%) and Curvibacter (21.24%). There were also genera such as Pelomonas. After the introduction of the pathogenic V. harveyi, the quantity of vibrios in the digestive tract increased from 7.2×105cfu·mL-1 to 10.9×105cfu·mL-1 as the stress time increased; the quantity of vibrio in the blood increased from 2×103cfu·mL-1 to 4.7×103cfu·mL-1; the quantity of vibrios in the foot muscle decreased after 48 h of V. harveyi stress and increased in the foot muscle at all other time. The Ace and Chao indices of the abalone digestive tract, gills, and foot muscles were lower than those of the control group at all time, except for the Ace and Chao indices of the abalone digestive tract after 24 h of V. harveyi stress and the Ace index of the digestive tract after 96 h of stress. The Shannon index was higher than the control in the digestive tract and foot at 24 h and in gill at 96h after V. harveyi infection and lower than the control at the remaining time. The introduction of V. harveyi increased the abundance of the Bacteroidetes as well as Fusobacteria in the digestive tract; the abundance of Proteobacteria in the gill decreased while that of Fusobacteria increased; the abundance of Proteobacteria in the foot muscles increased and that of Bacteroidetes decreased. At the genus level, the abundance of Pseudomonas, Bacteroides, and Propionibacterium increased and the abundance of Mycoplasma decreased in the digestive tract, except for a few time; the abundance of Bacteroides, Kordia, and Propionibacterium increased in gill; and the abundance of Pseudomonas increased in foot muscles. This study investigated the effect of the common pathogen V. harveyi on the quantity and the bacterial communities of the host flora from a microecological perspective and provided a new idea for further research on the response mechanism of the host to the pathogen V. harveyi and the prevention and control of vibriosis.
Keywords Vibrio harveyi; Haliotis discus hannai; bacterial diversity
ZHANG Ting , WANG Ruixuan , SUN Jingfeng , HUANG Jiajian , LIN Xiaozhi , ZHOU Fei , MA Xilan . Effect of Vibrio harveyi on the quantity and bacterial communities in different tissues of Haliotis discus hannai[J]. Journal of Tropical Oceanography, 2023 , 42(1) : 56 -65 . DOI: 10.11978/2022032
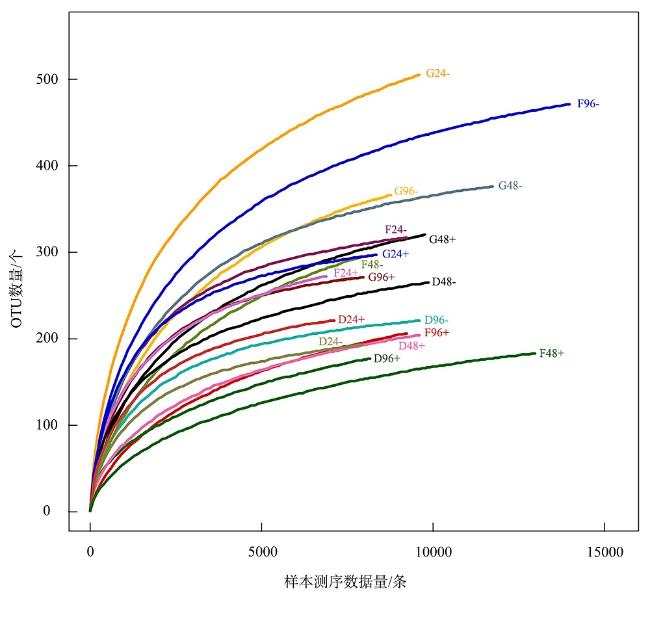
表1 对照组和试验组的微生物序列信息统计Tab. 1 Microbial sequence information statistics for control and experimental groups |
| 样本编号 | 有效序列数/条 | 优化序列数/条 | 可操作分类单位/个 |
|---|---|---|---|
| D24- | 8955 | 8306 | 7955 |
| D24+ | 9492 | 7336 | 7131 |
| D48- | 11704 | 10172 | 9873 |
| D48+ | 10900 | 9984 | 9621 |
| D96- | 10709 | 9890 | 9624 |
| D96+ | 9428 | 8599 | 8177 |
| G24- | 12658 | 10810 | 9628 |
| G24+ | 9801 | 8729 | 8353 |
| G48- | 13857 | 12173 | 11761 |
| G48+ | 12697 | 10108 | 9822 |
| G96- | 12527 | 9239 | 8785 |
| G96+ | 10654 | 8243 | 7986 |
| F24- | 11040 | 9683 | 9249 |
| F24+ | 8039 | 7348 | 6921 |
| F48- | 11463 | 8342 | 8063 |
| F48+ | 13942 | 13188 | 12983 |
| F96- | 16474 | 15156 | 13992 |
| F96+ | 13592 | 9453 | 9245 |
注: D表示消化道; G表示鳃; F表示足; -表示对照组; +表示试验组 |
表2 皱纹盘鲍消化道、鳃以及足部肌肉中细菌群落结构多样性指数Tab. 2 Structural diversity indices of bacterial communities in the digestive tract, gills and foot muscles of the H. discus hannai |
| 样本编号 | OTU | 覆盖率/% | Ace指数 | Chao指数 | Shannon指数 | Simpson指数 |
|---|---|---|---|---|---|---|
| D24- | 193 | 99.19 | 217 | 222 | 3.01 | 0.1077 |
| D24+ | 221 | 99.36 | 243 | 242 | 3.21 | 0.1326 |
| D48- | 265 | 99.22 | 313 | 326 | 3.68 | 0.0577 |
| D48+ | 204 | 99.24 | 260 | 246 | 2.71 | 0.1491 |
| D96- | 221 | 98.88 | 240 | 247 | 3.29 | 0.0878 |
| D96+ | 177 | 99.52 | 306 | 238 | 2.77 | 0.1333 |
| G24- | 505 | 99.39 | 606 | 621 | 4.56 | 0.0267 |
| G24+ | 297 | 99.52 | 319 | 330 | 3.93 | 0.0571 |
| G48- | 376 | 99.47 | 408 | 408 | 3.66 | 0.0823 |
| G48+ | 320 | 99.63 | 385 | 371 | 3.06 | 0.136 |
| G96- | 366 | 99.50 | 449 | 425 | 3.22 | 0.1544 |
| G96+ | 271 | 98.83 | 289 | 294 | 3.27 | 0.1294 |
| F24- | 317 | 98.68 | 342 | 350 | 3.4 | 0.1048 |
| F24+ | 272 | 99.11 | 312 | 326 | 3.97 | 0.0414 |
| F48- | 296 | 99.38 | 384 | 371 | 2.62 | 0.24 |
| F48+ | 183 | 99.48 | 246 | 240 | 1.33 | 0.5933 |
| F96- | 471 | 99.47 | 529 | 524 | 3.61 | 0.1473 |
| F96+ | 206 | 99.53 | 276 | 275 | 1.78 | 0.3866 |
注: D表示消化道; G表示鳃; F表示足; -表示对照组; +表示试验组 |
| [1] |
房沙沙, 林壮炳, 邱礽, 等, 2013. 粤东养殖区分离的2株海洋弧菌及其胞外产物对皱纹盘鲍致死毒性的初步分析[J]. 海洋科学, 37(8): 16-22.
|
| [2] |
郭战胜, 张海涛, 侯旭光, 2022. 4种鲍肠道菌群多样性比较[J]. 山东大学学报(理学版), 57(1): 1-7, 19.
|
| [3] |
韩敏敏, 李蜜, 刘昕明, 等, 2020. Khai岛和Pathiu岛珊瑚礁沉积物细菌多样性及细菌粗提物延缓秀丽隐杆线虫衰老活性研究[J]. 热带海洋学报, 39(5): 19-29.
|
| [4] |
蒋魁, 徐力文, 苏友禄, 等, 2016. 2012年-2014年南海海水养殖鱼类病原菌哈维弧菌分离株的耐药性分析[J]. 南方水产科学, 12(6): 99-107.
|
| [5] |
金珊, 王国良, 赵青松, 等, 2005. 海水网箱养殖大黄鱼弧菌病的流行病学研究[J]. 水产科学, 24(1): 17-19.
|
| [6] |
柯才焕, 2013. 我国鲍鱼养殖产业现状与展望[J]. 中国水产, (1): 27-30. (in Chinese)
|
| [7] |
李炳, 王瑞旋, 谢燕纯, 等, 2020. 广东沿海香港牡蛎消化道异养菌统计及其耐药性研究[J]. 海洋科学, 44(3): 50-58.
|
| [8] |
李沛翰, 林彦锋, 王凯英, 等, 2019. 16S扩增子技术研究海鲜样本菌群结构组成[J]. 军事医学, 43(4): 282-287.
|
| [9] |
林克冰, 吴建绍, 黄兆斌, 等, 2014. 一株斜带石斑鱼(Epinephelus coioides)病原菌的分离与鉴定[J]. 福建水产, 36(6): 419-427.
|
| [10] |
农业部渔业渔政管理局, 2014. 中国渔业统计年鉴2014[M]. 北京: 中国农业出版社: 23-42. (in Chinese)
|
| [11] |
孙慧敏, 戴世鲲, 王广华, 等, 2010. 南海北部巴士海峡深海沉积物中细菌多样性分析[J]. 热带海洋学报, 29(3): 41-46.
|
| [12] |
唐嘉威, 黄备, 孙沛雯, 等, 2020. 海洋保护区潮间带沉积物微生物群落的研究——以大连长海珍稀海洋生物保护区为例[J]. 海洋科学前沿, 7(1): 13-23.
|
| [13] |
王若璇, 2017. 鲍鱼养殖存在的主要问题与疾病综合防治对策[J]. 农业与技术, 37(4): 141. (in Chinese)
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}