
Journal of Tropical Oceanography >
Biodiversity, biogeography and seasonal variation of zooplankton Collodarians (Radiolaria) in surface waters from the northern Indian Ocean to the South China Sea*
Copy editor: YIN Bo
Received date: 2022-03-11
Revised date: 2022-04-24
Online published: 2022-04-26
Supported by
National Natural Science Foundation of China(41876207)
National Natural Science Foundation of China(42176080)
Chinese Academy of Sciences Project(QYZDY-SSW-DQC005)
Chinese Academy of Sciences Project(Y4SL021001)
Zooplankton collodarians with symbiontes play important roles in the organic carbon cycle and silicon cycle in the oligotrophic waters. However, studies on the collodiarian geographic distribution were few. In this study, the biodiversity, biogeography and seasonal variation of collodarians in the surface water of the northern Indian Ocean (NIO), the Malacca Strait (MLS) and the South China Sea (SCS) are revealed for the first time by using the ship-board plankton net to collect samples and using the Rose Bengal stains to distinguish between “living” and “dead” specimens. Totally, the 17 species of collodarians occurred in spring, with 27 species in winter. The collodarian diversity in NIO and MLS is generally less than that in SCS in spring, while in the winter, the former is higher than the latter, indicating that the regional biodiversity from NIO to the SCS is considerably affected by the East Asian monsoon. Moreover, the changes of collodarian community structure have biogeographic differences under the influence of East Asian monsoon. For example, the family Collosphaeridae is obviously dominant both in spring and winter, while the family Sphaerozoidae is dominant only in winter; the composition of dominant species is also different between the spring and the winter, suggesting the collodairian community in surface water is significantly affected by the seasonal changes. Under the influence of East Asian monsoon, the mixing of surface water enhanced, resulting in obvious changes in the compostion of collodarian species, which indicates that seasonal changes are the main factor that controls the community structure of collodarians in the study sea area. The abundance of collodarians is closely related to the regional environment. For instance, the collodarian abundance is very low in spring and winter from MLS to the Sunda Shelf, followed by SCS, and is high in NIO, reflecting their adaptability to specific marine environment. It is assumed that the influence of habitat on a large scale is higher than the control of monsoon change. Therefore, the biodiversity and abundance of collodarians can reflect the different ecological environmental signals, which further provides important observation data and a basic reference for the reconstruction of palaeooceanography and palaeoenvironment by using the collodarian substitution indexes in the future.
CHENG Xiawen , ZHANG Lanlan , QIU Zhuoya , XIANG Rong , CHANG Hu . Biodiversity, biogeography and seasonal variation of zooplankton Collodarians (Radiolaria) in surface waters from the northern Indian Ocean to the South China Sea*[J]. Journal of Tropical Oceanography, 2023 , 42(2) : 97 -112 . DOI: 10.11978/2022047
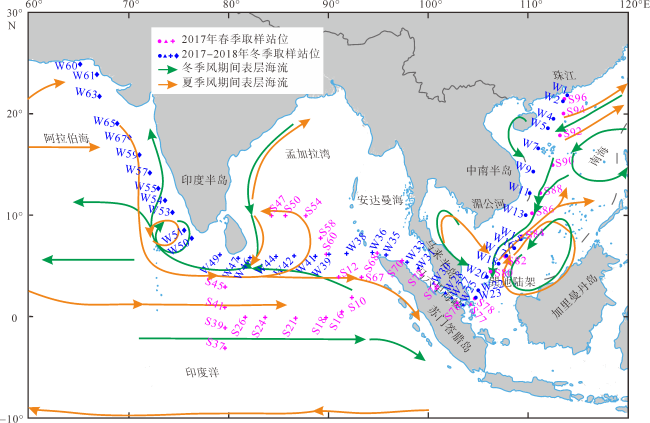
表1 走航水采集站位、经纬度、时间、样品流量以及胶体虫的物种数和总丰度Tab. 1 Sites, latitude, longitude, the time of sampling, sampling flow, number of species and total abundance of collodarian (specimens per one-meter square) |
| 站位 | 开始经度 | 开始纬度 | 结束经度 | 结束纬度 | 采样开始—结束时间 | 流量/L | 种数/个 | 总丰度/ (个·m-3) |
|---|---|---|---|---|---|---|---|---|
| S10 | 92°34′35″E | 1°59′58″N | 92°03′06″E | 3°00′02″N | 2017/03/14 00:25—05:40 | 6379 | 4 | 34 |
| S12 | 91°16′54″E | 3°58′35″N | 90°30′17″E | 4°59′58″N | 2017/03/15 04:15—09:25 | 5979 | 6 | 58 |
| S16 | 91°34′36″E | 0°33′31″N | 91°00′11″E | 0°00′09″N | 2017/03/16 22:30—00:30 (03/17) | 2991 | 3 | 5 |
| S18 | 90°00′44″E | 0°01′05″S | 88°59′57″E | 0°00′04″S | 2017/03/17 17:25—21:41 | 3291 | 3 | 5 |
| S21 | 87°00′40″E | 0°02′26″S | 86°00′05″E | 0°00′04″N | 2017/03/18 15:52—20:09 | 4626 | 4 | 15 |
| S24 | 83°58′53″E | 0°01′05″N | 83°00′11″E | 0°00′08″N | 2017/03/19 23:26—03:30 (03/19) | 6700 | 4 | 14 |
| S26 | 82°00′44″E | 0°02′27″N | 81°59′29″E | 0°59′58″S | 2017/03/20 16:39—00:17 (03/21) | 11242 | 6 | 18 |
| S37 | 80°00′40″E | 2°58′28″S | 80°00′05″E | 1°59′56″S | 2017/03/24 17:02—22:25 | 12169 | 9 | 234 |
| S39 | 80°00′06″E | 1°00′26″S | 79°54′59″E | 0°01′02″S | 2017/03/25 07:02—13:20 | 13029 | 6 | 23 |
| S41 | 79°59′38″E | 0°59′39″N | 79°59′57″E | 1°29′55″N | 2017/03/26 16:22—18:45 | 3309 | 6 | 48 |
| S45 | 80°01′06″E | 3°00′28″N | 79°00′13″E | 2°30′09″N | 2017/03/27 19:39—00:15 (03/28) | 6986 | 7 | 106 |
| S47 | 84°35′01″E | 10°00′25″N | 84°59′41″E | 10°00′02″N | 2017/04/05 02:00—05:00 | 4860 | 6 | 159 |
| S50 | 85°59′14″E | 10°00′5″N | 86°29′57″E | 10°00′03″N | 2017/04/05 16:30—21:50 | 9600 | 6 | 39 |
| S54 | 87°59′58″E | 10°00′00″N | 88°31′39″E | 10°00′04″N | 2017/04/07 04:06—07:05 | 5918 | 6 | 118 |
| S58 | 89°30′04″E | 7°48′26″N | 89°43′55″E | 7°02′44″N | 2017/04/08 13:45—16:55 | 5919 | 6 | 45 |
| S60 | 90°00′01″E | 6°14′16″N | 91°45′02″E | 1°16′07″N | 2017/04/09 23:04—18:50 (04/10) | 36886 | 5 | 33 |
| S67 | 93°31′21″E | 3°58′03″N | 94°57′12″E | 6°13′24″N | 2017/04/13 15:30—00:30 (04/14) | 5207 | 5 | 54 |
| S68 | 95°04′13″E | 6°19′35″N | 96°08′07″E | 6°01′58″N | 2017/04/14 02:00—06:00 | 3184 | 8 | 86 |
| S70 | 97°32′28″E | 5°34′59″N | 98°04′03″E | 5°23′52″N | 2017/04/14 11:15—13:10 | 1500 | 3 | 72 |
| S72 | 99°26′34″E | 4°11′17″N | 100°14′11″E | 3°28′15″N | 2017/04/14 19:36—23:50 | 3662 | 3 | 9 |
| S74 | 100°55′16″E | 2°52′51″N | 101°42′30″E | 2°20′29″N | 2017/04/15 03:26—07:11 | 4216 | 3 | 2 |
| S76 | 102°55′42″E | 1°32′43″N | 104°11′20″E | 1°15′11″N | 2017/04/15 12:16—17:50 | 5659 | 5 | 3 |
| S77 | 104°20′27″E | 1°17′36″N | 104°43′38″E | 1°51′45″N | 2017/04/15 18:26—21:12 | 5955 | 2 | 2 |
| S78 | 104°44′09″E | 1°53′41″N | 105°15′38″E | 3°24′24″N | 2017/04/15 21:20—03:31 (04/16) | 5104 | 0 | 0 |
| S80 | 106°19′34″E | 4°31′18″N | 107°17′53″E | 5°31′38″N | 2017/04/16 09:21—14:32 | 3817 | 5 | 2 |
| S82 | 107°44′55″E | 5°59′36″N | 108°42′21″E | 6°58′53″N | 2017/04/16 20:22—01:29 (04/17) | 5623 | 8 | 11 |
| S84 | 109°26′04″E | 8°00′14″N | 109°52′57″E | 9°01′21″N | 2017/04/17 06:12—10:15 | 6873 | 6 | 11 |
| S86 | 110°24′20″E | 10°15′55″N | 110°43′04″E | 10°59′28″N | 2017/04/17 14:59—17:51 | 5303 | 5 | 9 |
| S88 | 111°15′36″E | 12°15′26″N | 111°42′58″E | 13°19′07″N | 2017/04/17 22:41—02:41 (04/18) | 8553 | 6 | 29 |
| S90 | 112°27′19″E | 14°59′45″N | 112°55′57″E | 16°12′22″N | 2017/04/18 09:03—13:45 | 2081 | 9 | 26 |
| S92 | 113°12′14″E | 17°54′41″N | 113°25′45″E | 19°13′06″N | 2017/04/18 19:55—00:30 (04/19) | 4578 | 11 | 65 |
| S94 | 113°26′09″E | 19°15′31″N | 113°33′15″E | 19°59′14″N | 2017/04/19 06:02—09:12 | 3071 | 7 | 58 |
| S96 | 113°36′49″E | 21°29′10″N | 113°36′49″E | 21°29′10″N | 2017/04/19 12:10—14:55 | 5536 | 6 | 11 |
| W1 | 113°51′46″E | 21°49′33″N | 113°28′38″E | 21°16′53″N | 2017/12/31 13:21—15:48 | 6350 | 4 | 8 |
| W2 | 113°27′02″E | 21°14′30″N | 112°31′57″E | 20°29′54″N | 2017/12/31 15:57—19:13 | 8467 | 5 | 6 |
| W4 | 112°30′27″E | 19°30′42″N | 111°57′27″E | 18°35′28″N | 2017/12/31 23:16—03:10 (2018/01/01) | 10109 | 9 | 27 |
| W5 | 111°57′27″E | 18°35′28″N | 111°24′52″E | 17°41′14″N | 2018/01/01 03:17—06:57 | 9504 | 4 | 19 |
| W7 | 110°59′39″E | 16°36′21″N | 110°44′00″E | 15°31′08″N | 2018/01/01 11:17—15:26 | 10757 | 2 | 16 |
| W9 | 110°29′37″E | 14°19′18″N | 110°19′39″E | 13°23′27″N | 2018/01/01 19:43—23:01 | 8554 | 5 | 41 |
| W11 | 110°08′02″E | 12°12′26″N | 109°55′06″E | 11°02′54″N | 2018/01/02 03:15—07:25 | 10800 | 9 | 17 |
| W13 | 109°43′18″E | 10°02′36″N | 109°30′48″E | 8°59′25″N | 2018/01/02 11:03—14:49 | 9763 | 7 | 27 |
| W15 | 109°13′46″E | 7°41′43″N | 108°37′56″E | 6°49′44″N | 2018/01/02 19:32—23:16 | 9677 | 3 | 16 |
| W16 | 108°36′12″E | 6°48′02″N | 107°49′50″E | 6°03′43″N | 2018/01/02 23:24—03:15 (01/03) | 9979 | 3 | 14 |
| W17 | 107°49′10″E | 6°03′06″N | 107°01′02″E | 5°15′49″N | 2018/01/03 03:20—07:25 | 10584 | 1 | 2 |
| W18 | 107°01′08″E | 5°15′49″N | 106°22′11″E | 4°33′08″N | 2018/01/03 07:25—11:05 | 9504 | 3 | 4 |
| W20 | 105°50′16″E | 3°59′14″N | 105°18′10″E | 3°25′44″N | 2018/01/03 15:17—19:36 | 11189 | 1 | 0 |
| W22 | 105°00′30″E | 2°39′36″N | 104°45′49″E | 1°56′32″N | 2018/01/03 23:30—03:25 (01/04) | 10152 | 1 | 1 |
| W23 | 104°44′30″E | 1°55′37″N | 103°58′42″E | 1°14′59″N | 2018/01/04 03:35—08:15 | 12096 | 0 | 0 |
| W25 | 103°28′59″E | 1°13′27″N | 103°05′11″E | 1°32′23″N | 2018/01/04 10:10—12:08 | 5098 | 0 | 0 |
| W27 | 102°22′27″E | 1°53′19″N | 101°45′10″E | 2°22′26″N | 2018/01/04 15:00—17:39 | 6869 | 1 | 1 |
| W28 | 101°42′14″E | 2°24′50″N | 101°03′32″E | 2°47′35″N | 2018/01/04 17:52—20:47 | 7560 | 0 | 0 |
| W30 | 100°31′45″E | 3°20′55″N | 99°00′15″E | 4°51′22″N | 2018/01/04 23:30—02:45 (01/05) | 8424 | 1 | 6 |
| W31 | 99°00′15″E | 4°51′22″N | 98°35′31″E | 5°09′28″N | 2018/01/05 07:49—11:15 | 8899 | 2 | 3 |
| W33 | 97°53′43″E | 5°26′39″N | 96°59′05″E | 5°45′41″N | 2018/01/05 15:09—18:35 | 8813 | 1 | 6 |
| W35 | 95°48′01″E | 6°08′43″N | 94°24′58″E | 6°22′20″N | 2018/01/05 23:10—04:10 (01/06) | 12960 | 7 | 89 |
| W36 | 94°24′20″E | 6°22′05″N | 93°10′42″E | 6°20′14″N | 2018/01/06 04:15—07:25 | 8208 | 8 | 187 |
| W38 | 91°50′12″E | 6°17′01″N | 91°09′49″E | 6°13′44″N | 2018/01/06 12:05—15:33 | 8986 | 3 | 19 |
| W39 | 90°05′46″E | 6°13′38″N | 89°53′13″E | 6°12′08″N | 2018/01/06 15:47—20:07 | 11232 | 7 | 53 |
| W41 | 88°43′20″E | 6°00′02″N | 86°36′34″E | 6°03′36″N | 2018/01/07 00:30—07:00 | 16848 | 7 | 53 |
| W42 | 86°36′34″E | 6°03′36″N | 85°24′59″E | 6°00′39″N | 2018/01/07 07:00—11:20 | 11232 | 8 | 47 |
| W44 | 84°48′26″E | 5°59′12″N | 83°32′53″E | 5°56′08″N | 2018/01/07 14:40—19:18 | 12010 | 7 | 23 |
| W46 | 82°11′45″E | 5°52′11″N | 81°01′51″E | 5°49′35″N | 2018/01/08 00:20—04:11 | 9979 | 9 | 47 |
| W47 | 81°01′11″E | 5°49′30″N | 80°08′51″E | 5°50′29″N | 2018/01/08 04:20—07:25 | 7992 | 7 | 26 |
| W49 | 79°12′20″E | 6°10′02″N | 76°23′57″E | 7°46′30″N | 2018/01/08 11:20—23:30 | 31536 | 11 | 64 |
| W50 | 76°23′45″E | 7°46′55″N | 75°38′12″E | 8°32′19″N | 2018/01/08 23:40—03:35 (01/09) | 10152 | 11 | 212 |
| W51 | 75°37′44″E | 8°33′10″N | 75°00′04″E | 9°22′07″N | 2018/01/09 03:45—07:15 | 9072 | 10 | 109 |
| W53 | 74°28′16″E | 10°19′31″N | 73°48′23″E | 11°19′06″N | 2018/01/09 11:30—15:56 | 11491 | 15 | 201 |
| W54 | 73°42′28″E | 11°29′44″N | 73°03′41″E | 12°38′17″N | 2018/01/09 16:40—21:29 | 12485 | 12 | 317 |
| W55 | 73°02′35″E | 12°40′36″N | 72°38′59″E | 13°28′04″N | 2018/01/09 21:39—00:59 (01/10) | 8640 | 10 | 309 |
| W57 | 72°12′13″E | 14°11′45″N | 71°43′50″E | 14°59′05″N | 2018/01/10 03:55—07:10 | 8424 | 16 | 120 |
| W59 | 71°08′52″E | 15°58′47″N | 70°35′10″E | 16°59′41″N | 2018/01/10 11:20—15:42 | 11318 | 11 | 193 |
| W60 | 65°14′32″E | 24°54′00″N | 65°15′03″E | 24°50′35″N | 2018/02/02 16:25—20:05 | 9504 | 5 | 13 |
| W61 | 66°55′35″E | 23°51′08″N | 66°57′57″E | 22°47′34″N | 2018/02/04 21:20—01:20 (02/05) | 10368 | 3 | 3 |
| W63 | 67°11′19″E | 21°42′47″N | 68°04′34″E | 20°32′54″N | 2018/02/05 05:25—10:30 | 13176 | 6 | 9 |
| W65 | 68°56′39″E | 19°03′34″N | 69°28′14″E | 18°37′51″N | 2018/02/05 16:04—19:40 | 9331 | 9 | 56 |
| W67 | 70°10′07″E | 17°41′06″N | 70°42′41″E | 16°45′22″N | 2018/02/05 23:59—03:53 | 10109 | 6 | 66 |
图2 研究样品采集期间的水文环境a. 冬季海表温度; b. 春季海表温度; c. 冬季海表盐度; d. 春季海表盐度; e. 冬季硅酸盐浓度; f. 春季硅酸盐浓度; g. 冬季磷酸盐浓度; h. 春季硅酸盐浓度。该图基于国家测绘地理信息局标准地图服务网站下载的审图号为GS(2016)1663的标准地图制作, 底图无修改 Fig. 2 Hydrological environment during the sampling periods. (a) sea surface temperature in winter; (b) sea surface temperature in spring; (c) sea surface salinity in winter; (d) sea surface salinity in spring; (e) silicate concentration in winter; (f) silicate concentration in spring; (g) phosphate concentration in winte; (h) phosphate concentration in spring |
图3 胶体虫在冬季和春季的物种数和多样性变化a. 冬季物种数; b. 春季物种数; c. 冬季多样性; d. 春季多样性。该图基于国家测绘地理信息局标准地图服务网站下载的审图号为GS(2016)1663的标准地图制作, 底图无修改 Fig. 3 Variation in species number and diversity of collodarians in winter and spring. (a) species number in winter; (b) species number in spring; (c) diversity in winter; (d) diversity in spring |
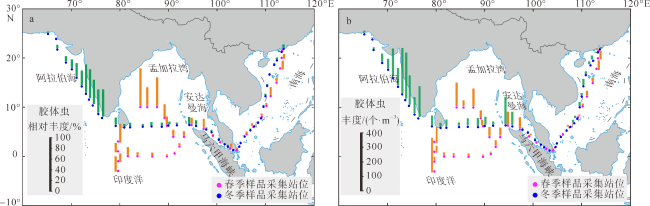
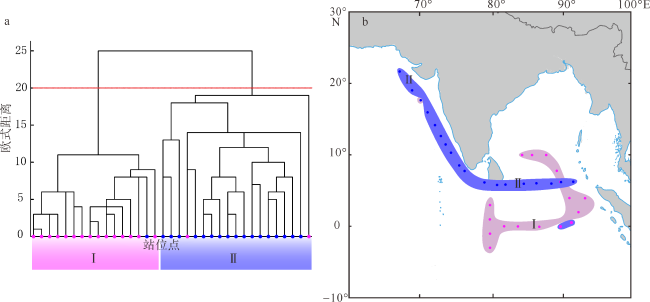
图4 北印度洋—南海春季和冬季胶体虫的相对丰度(a)和丰度(b)变化马六甲海峡部分站位无胶体虫。该图基于国家测绘地理信息局标准地图服务网站下载的审图号为GS(2016)1663的标准地图制作, 底图无修改 Fig. 4 Relative abundance (a) and absolute abundance (b) of Collodaria in spring and winter in the northern Indian Ocean-South China Sea. There is no collodarian observed at sites W23, W25 and W28 in the MLS |
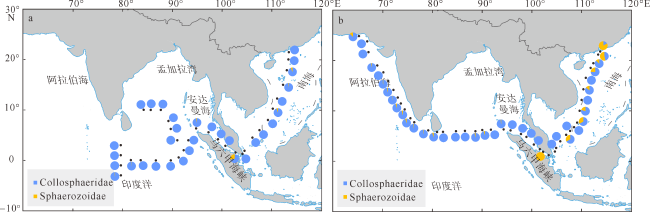
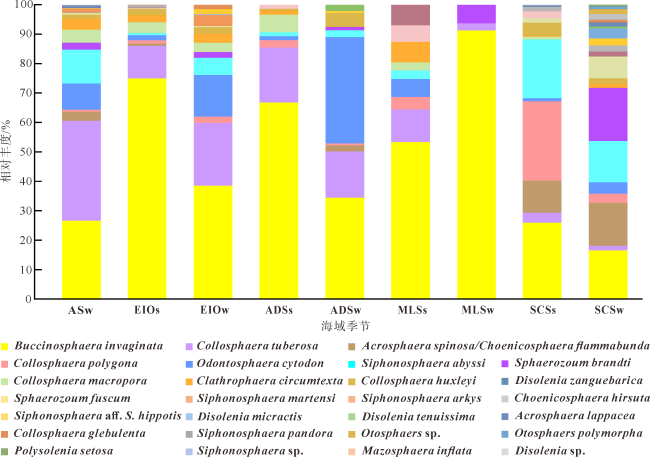
图6 不同海域春季和冬季胶体虫的物种组成变化ASw: 阿拉伯海冬季; EIOs: 东印度洋春季; EIOw: 东印度洋冬季; ADSs: 安达曼海春季; ADSw: 安达曼海冬季; MLSs; 马六甲海峡春季; MLSw: 马六甲海峡冬季; SCSs: 南海春季; SCSw: 南海冬季 Fig. 6 Composition of collodarian species in spring and winter in each sea areas. ASw: Arabian Sea in winter; EIOs: eastern Indian Ocean in spring; EIOw: eastern Indian Ocean in winter; ADSs: Andaman Sea in spring; ADSw: Andaman Sea in winter; MLSs: Malacca Strait in spring; MLSw: Malacca Strait in winter; SCSs: south China sea in spring; SCSw: south China sea in winter |
表2 研究区胶体虫物种在春季和冬季表层水体中的优势度(Y)Tab. 2 Dominance degree of collodarian species in spring and winter surface water in the studied areas |
| 胶体虫种名 | 春季优势度 | 冬季优势度 |
|---|---|---|
| Acrosphaera lappacea (Haeckel) | 0.0001 | 0.0009 |
| Acrosphaera spinosa (Haeckel) / Choenicosphaera flammabunda Haeckel | 0.0069 | 0.0104 |
| Buccinosphaera invaginata Haeckel | 0.6214 | 0.2210 |
| Choenicosphaera hirsuta (Ehrenberg) | 0.0010 | 0.0002 |
| Clathrophaera circumtexta Haeckel | 0.0054 | 0.0075 |
| Collosphaera glebulenta Bjørklund & Goll | 0.0000 | 0.0002 |
| Collosphaera huxleyi Müller | 0.0092 | 0.0075 |
| Collosphaera macropora Popofsky | 0.0192 | 0.0116 |
| Collosphaera polygona Haeckel | 0.0296 | 0.0027 |
| Collosphaera tuberosa Haeckel | 0.0904 | 0.1523 |
| Disolenia micractis (Ehrenberg) | 0 | 0.0001 |
| Disolenia sp. | 0 | 0.0000 |
| Disolenia tenuissima (Hilmers) | 0 | 0.0002 |
| Disolenia zanguebarica Ehrenberg | 0.0005 | 0.0023 |
| Mazosphaera inflata (Haeckel) | 0 | 0.0000 |
| Odontosphaera cyrtodon Haeckel | 0.0058 | 0.0660 |
| Otosphaera polymorpha Haeckel | 0 | 0.0000 |
| Otosphaers sp. | 0 | 0.0000 |
| Polysolenia setosa Ehrenberg | 0 | 0.0000 |
| Siphonosphaera abyssi (Ehrenberg) | 0.0188 | 0.0480 |
| Siphonosphaera aff. S. hippotis (Haeckel) | 0 | 0.0015 |
| Siphonosphaera arkys Su | 0 | 0.0032 |
| Siphonosphaera martensi Brandt | 0.0001 | 0.0001 |
| Siphonosphaera pandora (Haeckel) | 0.0000 | 0.0001 |
| Siphonosphaera sp. | 0 | 0.0000 |
| Sphaerozoum brandti Popofsky | 0 | 0.0183 |
| Sphaerozoum fuscum Meyen | 0.0015 | 0 |
注: 加粗表示Y > 0.02 |
图7 胶体虫优势种在春、冬季的绝对丰度和相对丰度分布a1 ~ a4: Buccinosphaera invaginata; b1 ~ b4: Collosphaera tuberosa; c1 ~ c4: Odontosphaera cyrtodon; d1 ~ d4: Siphonosphaera abyssi; e1 ~ e4: Collosphaera polygona。该图基于国家测绘地理信息局标准地图服务网站下载的审图号为GS(2016)1663的标准地图制作, 底图无修改 Fig. 7 Absolute and relative abundances of dominant collodarian species in spring and winter, a1~a4: Buccinosphaera invaginata; b1~b4: Collosphaera tuberosa; c1~c4: Odontosphaera cyrtodon; d1~d4: Siphonosphaera abyssi; e1~e4: Collosphaera polygona |
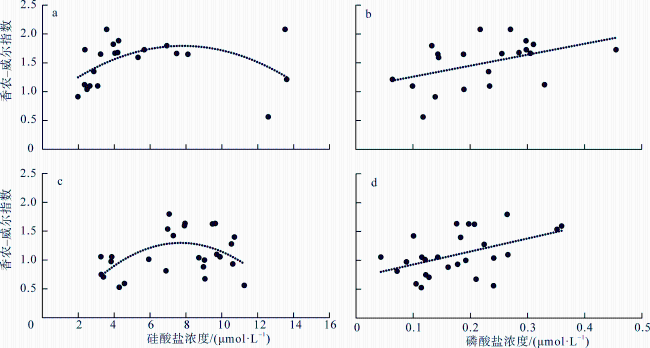
图9 南海、北印度洋胶体虫多样性与采样站环境变量之间的相关性a. 冬季胶体虫多样性与硅酸盐浓度的关系; b. 春季胶体虫多样性与硅酸盐浓度的关系; c. 冬季胶体虫多样性与磷酸盐浓度的关系; d. 春季胶体虫浓度与磷酸盐浓度的关系。图中黑色圆点为实际数据, 虚线为拟合线 Fig. 9 Correlations between the collodarian diversity and mean environment variables across the sampling sites. (a) relation with the silicate concentration in winter; (b) relation with the silicate concentration in spring; (c) relation with the phosphate concentration in winter; (d) relation with the phosphate concentration in spring |
| [1] |
陈木宏, 谭智源, 1996. 南海中、北部沉积物中的放射虫[M]. 北京: 科学出版社:1-271.
|
| [2] |
范锦晓, 胡松, 2020. 潮汐和风对马六甲海峡海流的影响[J]. 热带海洋学报, 39(4): 13-24.
|
| [3] |
胡维芬, 张兰兰, 陈木宏, 等, 2015. 南海断面春季活体放射虫生态分布及其对环境的响应[J]. 中国科学: 地球科学, 45(1): 83-98.
|
| [4] |
黄锦平, 吴泽涛, 苏玉萍, 等, 2022. 不同浓度和形态磷模拟调控浮游植物群落演替实验[J]. 环境科学学报, 42(2): 422-429.
|
| [5] |
刘颖, 严幼芳, 凌征, 2020. 北印度洋障碍层厚度气候态和季节变化特征及其成因初步分析[J]. 热带海洋学报, 39(5): 98-108.
|
| [6] |
沈国英, 施并章, 2002. 海洋生态学[M]. 北京: 海洋生态学:158-159.
|
| [7] |
斯特列尔科夫 A A, 列雪特妮阿克 V V, 1962. 中国南海海南岛南端地区的群体放射虫[J]. 海洋科学集刊, 1: 121-139.
|
| [8] |
谭智源, 1998. 中国动物志[M]. 北京: 科学出版社:1-360.
|
| [9] |
谭智源, 陈木宏, 1999. 中国近海的放射虫[M]. 北京: 科学出版社:1-404.
|
| [10] |
王磊, 冷晓云, 孙庆杨, 等, 2015. 春季季风间期巽他陆架和马六甲海峡表层海水浮游植物群落结构研究[J]. 海洋学报, 37(2): 120-129.
|
| [11] |
王雪松, 陈忠, 许安涛, 等, 2022. 南海东北部深海盆末次冰盛期以来陆源碎屑粒度特征及影响因素[J]. 热带海洋学报, 41(1): 158-170.
|
| [12] |
俞慕耕, 1987. 略论马六甲海峡的水文特点[J]. 海洋湖沼通报, (2): 6-16.
|
| [13] |
张杰, 张兰兰, 陈木宏, 等, 2020. 现代放射虫的高阶分类现状及其生态学意义[J]. 微体古生物学报, 37(1): 82-98.
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
/
| 〈 |
|
〉 |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}