
Journal of Tropical Oceanography >
Study on the effect of Oyster hydrolysates on improving Lactation function in Postpartum hypogalactism
Copy editor: YIN Bo
Received date: 2022-04-11
Revised date: 2022-06-17
Online published: 2022-06-30
Supported by
China Agricultural Research System of MOF and MARA(CARS-49)
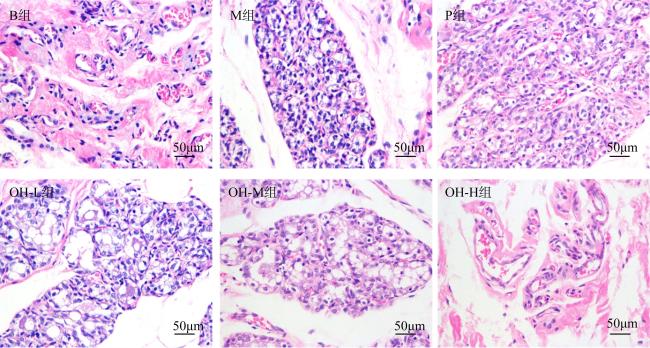
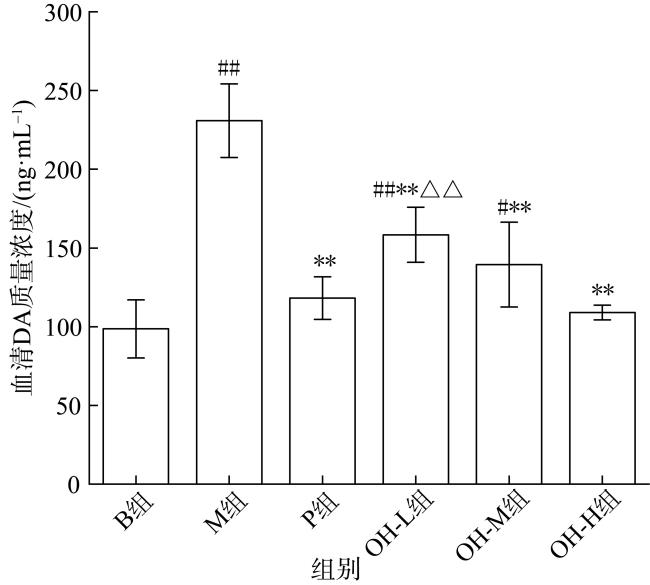
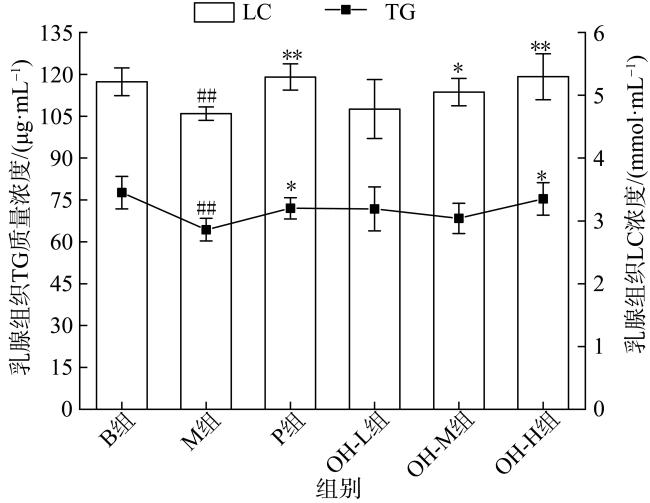
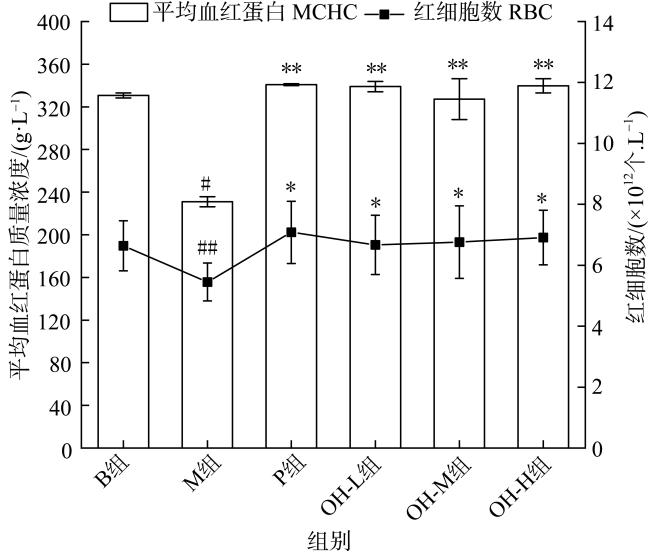
In this paper, oyster hydrolysis products were prepared from Crassostrea hongkongensis as raw materials, and the basic nutrition and amino acid composition of oyster hydrolysis products were analyzed, and the improvement effect of oyster hydrolysis products on the lactation function of rats with postpartum lactation caused by Bromocriptine mesylate was explored. The results showed that protein content of oyster hydrolysis products was higher, which is followed by sugar, fat content was lower, and peptide content was 33.4%. The essential amino acids in oyster hydrolysis products accounted for 39.8% of the total amino acids and contained a high proportion of branched amino acids. The results of animal experiment showed that compared with model group, the lactation volume of rats in oyster hydrolysis products of high-dose group was significantly increased (p < 0.05), which was better than that in the positive control group. The index of mammary tissue and organ in each oyster hydrolysis products dose group was significantly increased (p < 0.01), which was close to the positive control group, but did not return to normal level. HE staining showed that there were more acinus in mammary gland tissues in all oyster hydrolysis products dose groups, which were round and full with more secretions, and the acinus had obvious enlargement and hyperplasia. Compared with the model group, the acinus had obvious recovery. Compared with model group, the contents of prolactin (PRL) and prolactin receptor (PRLR) in serum and mammary tissue of oyster hydrolysis products groups were largely increased (p < 0.05), indicating that oyster hydrolysis products could promote the secretion of prolactin in lactation-deficient rats, while the content of serum dopamine (DA) was greatly decreased (p < 0.01). These results showed that oyster hydrolysis products can reduce the high level of dopamine induced by bromocystine. In addition, oyster hydrolysis products strongly increased the contents of lactose (LC) and triglyceride (TG) in breast tissue (p < 0.05), and improved the nutritional composition of milk. The average hemoglobin (MCHC) and red blood cell (RBC) contents in all oyster hydrolysis products dose groups were also greatly increased (p < 0.05), which had a certain improvement effect on postpartum deficiency of qi and blood. Conclusion: Oyster hydrolysis products can improve the symptoms of lactation deficiency and increase milk yield after delivery, which has the potential to promote lactation.
WEI Xue , QIN Xiaoming , CHEN Suhua , ZHANG Kaijia , LIN Haisheng , ZHENG Huina , GAO Jialong . Study on the effect of Oyster hydrolysates on improving Lactation function in Postpartum hypogalactism[J]. Journal of Tropical Oceanography, 2023 , 42(2) : 141 -152 . DOI: 10.11978/2022075
表1 动物分组及处理Tab. 1 Grouping and treatment of animals |
| 组别 | 母鼠数量/只 | 每窝仔鼠数量/只 | 处理方式 |
|---|---|---|---|
| 空白组(B组) | 6 | 12 | 用1mL·100g-1蒸馏水灌胃, 每天一次 |
| 模型组(M组) | 6 | 12 | 同时用甲磺酸溴隐亭溶液(浓度0.5mg·mL-1、体积1mL·100g-1)和1mL·100g-1蒸馏水灌胃, 每天一次 |
| 阳性药物组(P组) | 6 | 12 | 同时用甲磺酸溴隐亭溶液(浓度0.5mg·mL-1、体积1mL·100g-1)和补血生乳颗粒(浓度267mg·mL-1、体积1mL·100g-1)灌胃, 每天一次 |
| 牡蛎酶解产物低剂量组(OH-L组) | 6 | 12 | 同时用甲磺酸溴隐亭溶液(浓度0.5mg·mL-1、体积1mL·100g-1)和牡蛎酶解粉(浓度31.25mg·mL-1、体积1mL·100g-1)灌胃, 每天一次 |
| 牡蛎酶解产物中剂量组(OH-M 组) | 6 | 12 | 同时用甲磺酸溴隐亭溶液(浓度0.5mg·mL-1、体积1L·100g-1)和牡蛎酶解粉(浓度62.5mg·mL-1、体积1 mL·100g-1)灌胃, 每天一次 |
| 牡蛎酶解产物高剂量组(OH-H 组) | 6 | 12 | 同时用甲磺酸溴隐亭溶液(浓度0.5mg·mL-1、体积1mL·100g-1)和牡蛎酶解粉(浓度125mg·mL-1、体积1mL·100g-1)灌胃, 每天一次 |
表2 牡蛎酶解产物(OH)基本营养成分组成( , n=3 )Tab. 2 Basic nutritional components of OH ( , n=3 ) |
| 成分 | 肽/% | 水分/% | 脂肪/% | 总糖/% | 蛋白质/% | 灰分/% |
|---|---|---|---|---|---|---|
| OH | 33.4±0.14 | 7.17±0.15 | 4.55±0.21 | 33.83±1.52 | 45.09±0.98 | 6.60±0.17 |
表3 OH氨基酸组成表Tab. 3 Amino acid composition of OH |
| 种类 | 水解氨基酸含量/(g·100g-1) | 游离氨基酸含量/(g·100g-1) |
|---|---|---|
| 天门冬氨酸(ASP) | 3.81 | 0.74 |
| 苏氨酸(THR)* | 1.67 | 1.37 |
| 丝氨酸(SER) | 1.80 | 1.00 |
| 谷氨酸(GLU) | 5.94 | 1.97 |
| 脯氨酸(PRO) | 1.46 | 0.59 |
| 甘氨酸(GLY) | 2.45 | 0.65 |
| 丙氨酸(ALA) | 2.20 | 1.56 |
| 缬氨酸(VAL)* | 2.16 | 1.30 |
| 蛋氨酸(MET)* | 1.34 | 0.97 |
| 异亮氨酸(ILE)* | 1.79 | 1.43 |
| 亮氨酸(LEU)* | 2.79 | 2.39 |
| 酪氨酸 TYR | 1.04 | 2.09 |
| 苯丙氨酸(PHE)* | 1.57 | 2.02 |
| 赖氨酸(LYS)* | 2.84 | 0.92 |
| 组氨酸(HIS) | 0.48 | 0.27 |
| 精氨酸(ARG) | 2.31 | 1.52 |
| 氨基酸总和(TAA) | 35.60 | 20.80 |
| 必需氨基酸(EAA) | 14.16 | 10.40 |
| 非必需氨基酸(NEAA) | 21.44 | 10.40 |
| EAA与TAA比值/% | 39.80 | 50.00 |
| EAA与NEAA比值/% | 66.00 | 100.00 |
注: *为必需氨基酸 |
表4 乳腺器官指数( , n=6 )Tab. 4 Mammary organ index ( , n=6 ) |
| 组别 | B组 | M组 | P组 | OH-L组 | OH-M组 | OH-H组 |
|---|---|---|---|---|---|---|
| 指数 | 6.49±0.72 | 3.35±0.3## | 5.65±0.69** | 4.76±0.64##** | 4.81±0.57##** | 4.98±0.44##** |
注: 与B组比较, #表示差异显著(p < 0.05), ##表示差异极显著(p < 0.01); 与M组比较, *表示差异显著(p < 0.05), **表示差异极显著(p < 0.01); 与P组比较, △表示差异显著(p < 0.05), △△表示差异极显著(p < 0.01) |
表5 脏器器官指数( , n=6 )Tab. 5 Organ index ( , n=6 ) |
| 指标 | B组 | M组 | P组 | OH-L组 | OH-M组 | OH-H组 |
|---|---|---|---|---|---|---|
| 心脏指数 | 0.45±0.11 | 0.43±0.03 | 0.43±0.05 | 0.40±0.03 | 0.44±0.05 | 0.43±0.06 |
| 胸腺指数 | 0.09±0.02 | 0.11±0.02 | 0.12±0.01 | 0.11±0.01 | 0.11±0.02 | 0.12±0.02 |
| 脾脏指数 | 0.22±0.01 | 0.24±0.02 | 0.24±0.02 | 0.23±0.01 | 0.25±0.02 | 0.25±0.04 |
| 肝脏指数 | 3.94±0.42 | 3.41±0.37 | 3.20±0.22 | 3.20±0.24 | 3.28±0.21 | 3.41±0.21 |
| 肾脏指数 | 0.70±0.07 | 0.74±0.07 | 0.74±0.04 | 0.75±0.08 | 0.72±0.06 | 0.71±0.05 |
表6 每小时泌乳量( , n=6 )(单位: g·h-1 )Tab. 6 Lactating capacity per hour ( , n=6) |
| 时间 | B组 | M组 | P组 | OH-L组 | OH-M组 | OH-H组 |
|---|---|---|---|---|---|---|
| 第1天 | 2.28±0.73 | 1.49±0.40## | 3.35±0.50** | 1.75±1.94 | 2.51±0.48** | 2.93±0.50** |
| 第2天 | 2.29±1.04 | 2.57±1.04 | 2.07±0.86 | 0.81±1.82 | 1.21±0.41 | 3.19±0.32 |
| 第3天 | 4.11±0.79 | 1.98±1.26# | 1.60±0.77 | 2.33±0.77 | 0.89±0.83 | 2.02±0.57 |
| 第4天 | 4.46±0.56 | 0.90±1.13## | 2.17±0.77 | 0.71±1.42 | 1.99±0.88 | 1.74±0.12 |
| 第5天 | 6.38±1.15 | 0.44±0.48## | 1.05±0.98 | 1.77±1.24 | 0.67±0.41 | 2.47±1.00** |
| 第6天 | 6.20±1.1 | 0.03±0.48## | 1.82±0.31** | 1.05±1.39 | 0.13±0.66 | 2.79±0.89** |
| 第7天 | 4.88±1.62 | 0.39±0.51## | 0.05±0.79 | 0.57±1.29 | 1.00±0.79 | 3.10±0.85** |
| 第8天 | 4.74±1.45 | 0.05±0.10## | 1.64±2.10** | 0.93±1.48 | 0.18±0.23 | 1.11±0.70 |
注: 与B组比较, #表示差异显著(p < 0.05), ##表示差异极显著(p < 0.01); 与M组比较, *表示差异显著( p < 0.05), **表示差异极显著(p < 0.01); 与P组比较, △表示差异显著(p < 0.05), △△表示差异极显著(p < 0.01) |
表7 泌乳量总和平均值( , n=6 )(单位: g )Tab. 7 Average total milk yield ( , n=6) |
| 组别 | B组 | M组 | P组 | OH-L组 | OH-M组 | OH-H组 |
|---|---|---|---|---|---|---|
| 平均值 | 4.39±1.81 | 0.82±1.30##△△ | 1.72±1.35##** | 1.16±1.65## | 1.08±1.09##△ | 2.42±0.97##**△ |
注: 与B组比较, #表示差异显著(p < 0.05), ##表示差异极显著(p < 0.01); 与M组比较, *表示差异显著(p < 0.05), **表示差异极显著(p < 0.01); 与P组比较, △表示差异显著(p < 0.05), △△表示差异极显著(p < 0.01) |
表8 仔鼠存活率( , n=12 ) (单位: %)Tab. 8 Survival rate of their offspring ( , n=12) |
| 组别 | 第3天 | 第4天 | 第5天 | 第6天 | 第7天 | 第8天 |
|---|---|---|---|---|---|---|
| B组 | 100±0.00 | 100±0.00 | 100±0.00 | 100±0.00 | 100±0.00 | 100±0.00 |
| M组 | 100±0.00 | 100±0.00 | 94.44±8.61 | 88.88±6.21## | 84.72±3.40## | 77.78±4.30## |
| P组 | 100±0.00 | 100±0.00 | 100±0.00 | 98.61±3.40* | 95.84±4.56** | 87.50±4.57** |
| OH-L组 | 100±0.00 | 97.22±4.30 | 98.61±3.40 | 98.61±3.40* | 87.50±4.57##△△ | 81.94±3.40△ |
| OH-M组 | 100±0.00 | 100±0.00 | 98.61±3.40 | 95.84±4.56 | 94.45±4.30** | 84.72±3.40** |
| OH-H组 | 100±0.00 | 100±0.00 | 100±0.00 | 100±0.00** | 98.61±3.40** | 93.06±3.40**△ |
注: 与B组比较, #表示差异显著(p < 0.05), ##表示差异极显著(p < 0.01); 与M组比较, *表示差异显著(p < 0.05), **表示差异极显著(p < 0.01); 与P组比较, △表示差异显著(p < 0.05), △△表示差异极显著(p < 0.01) |
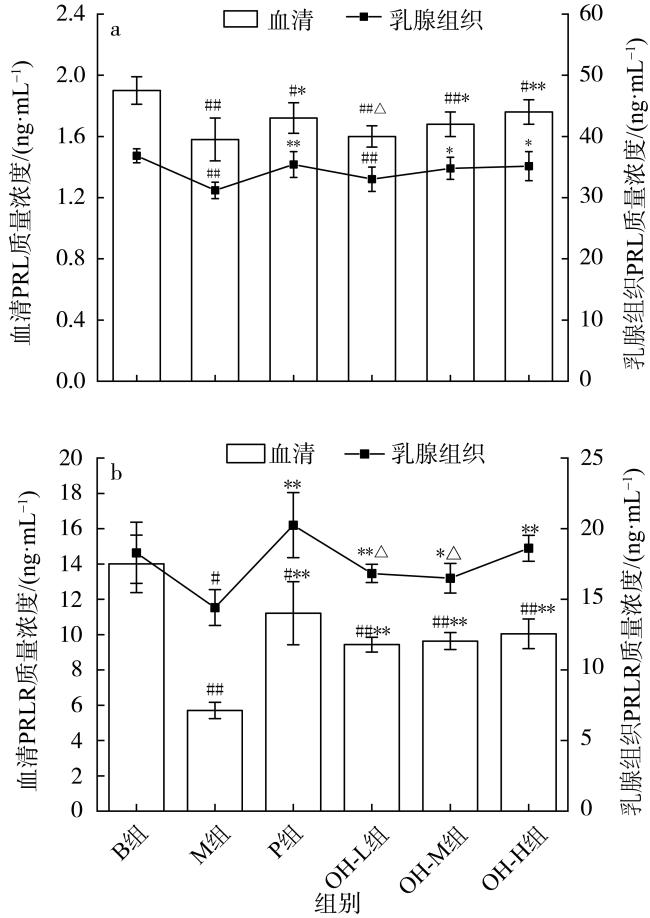
图2 缺乳大鼠血清和乳腺组织催乳素(PRL, a)、催乳素受体(PRLR, b)的质量浓度 ( , n=6 )与B组比较, #表示差异显著(p < 0.05), ##表示差异极显著(p < 0.01); 与M组比较, *表示差异显著(p < 0.05), **表示差异极显著(p < 0.01); 与P组比较, △表示差异显著(p < 0.05), △△表示差异极显著(p < 0.01) Fig. 2 Content of PRL (a) and PRLR (b) in mammary gland and serum of postpartum lactation deficient rats ( , n=6) |
| [1] |
谌素华, 秦小明, 章超桦, 等, 2021. 牡蛎酶解产物对超负荷哺乳大鼠泌乳的影响[J]. 南方水产科学, 17(6): 107-114.
|
| [2] |
底爱英, 高文生, 2013. 溴隐亭治疗垂体泌乳素腺瘤的研究进展[J]. 脑与神经疾病杂志, 21(1): 78-80.
|
| [3] |
董世起, 2015. 催乳汤对产后缺乳模型大鼠的治疗作用及其机制研究[D]. 北京: 中国农业大学:103.
|
| [4] |
付琳, 张丽, 王高富, 等, 2020. 小肽调控奶牛乳腺乳蛋白合成的作用机制[J]. 动物营养学报, 32(5): 1989-1996.
|
| [5] |
高亚红, 2016. 产妇泌乳以及产后康复中应用补气益血组方干预的临床价值分析[J]. 当代医学, 22(36): 175-176.
|
| [6] |
谷子林, 刘亚娟, 陈赛娟, 等, 2020. 复方中药小肽对母兔泌乳性能及配种受胎率的影响[J]. 中国兽医学报, 40(11): 2246-2250.
|
| [7] |
农业农村部渔业渔政管理局, 全国水产技术推广总站, 中国水产学会, 2021. 中国渔业统计年鉴[M]. 北京: 中国农业出版社.
FISHERY ADMINISTRATION BUREAU OF MINISTRY OF AGRICULTURE AND RURAL AFFAIRS, NATIONAL FISHERIES TECHNOLOGY EXTENSION CENTER, CHINA SOCIETY OF FISHERIES, 2021. China fisheries statistical yearbook[M]. Beijing: China Agriculture Press. (in Chinese with English abstract)
|
| [8] |
胡培丽, 梁怿, 林飞, 2014. 鱼牡蛎生乳灵胶囊促进大鼠泌乳功能的实验研究[J]. 癌变·畸变·突变, 26(4): 285-287.
|
| [9] |
蒋博, 汪金林, 王海燕, 等, 2022. 鲟鱼头汤对产后缺乳大鼠泌乳功能的促进作用[J/OL]. 食品科学, 1-12.
|
| [10] |
李君, 杨文卓, 候霞飞, 等, 2019. 油酸和亚油酸对山羊乳腺细胞甘油三酯含量及乳脂合成相关基因表达的影响[J]. 中国畜牧兽医, 46(2): 380-386.
|
| [11] |
李婉, 曹文红, 章超桦, 等, 2017. 牡蛎酶解产物的组成特点及其体外免疫活性[J]. 食品工业科技, 38(16): 35-42.
|
| [12] |
林超, 2020. 浅析我国婴儿喂养的现状和应对策略[J]. 决策探索(下), 12: 91-92.
|
| [13] |
林国兴, 刘志勇, 李芳, 2021. 中医药膳治疗产后乳少的研究现状[J]. 江西中医药, 52(2): 78-80.
|
| [14] |
林怀美, 陈淑音, 林跃进, 等, 2021. 母乳成分分析与乳母膳食的相关性研究[J]. 基层医学论坛, 25(29): 4147-4149.
|
| [15] |
林曼, 2018. 槲皮素对产后缺乳模型小鼠的治疗作用及其机制研究[D]. 广东: 广东药科大学.
|
| [16] |
孙龙生, 魏凯, 安振华, 等, 2011. 水产动物理想蛋白质模式研究进展[J]. 动物营养学报, 23(9): 1453-1458.
|
| [17] |
王一飞, 宋阳, 陈宝艳, 等, 2018. “米酒鸡”食疗方对产后缺乳母鼠催乳素受体及β酪蛋白表达的影响[J]. 广州中医药大学学报, 35(4): 689-693.
|
| [18] |
吴缃琦, 孙晓勉, 何珊茹, 2017. 6个月内婴儿纯母乳喂养现状及影响因素分析[J]. 中国儿童保健杂志, 25(1): 90-93.
|
| [19] |
吴园涛, 任小波, 孙恢礼, 2012. 关于我国海洋生物资源领域科技布局的若干思考[J]. 热带海洋学报, 31(1): 79-84.
|
| [20] |
许丹, 林峰, 朱小语, 等, 2016. 牡蛎肽对免疫抑制小鼠免疫功能的影响[J]. 北京大学学报(医学版), 48(03): 392-397.
|
| [21] |
杨发明, 林海生, 秦小明, 等, 2020. 牡蛎酶解产物促进皮肤软组织创伤愈合的作用[J]. 上海海洋大学学报, 29(4): 632-640.
|
| [22] |
杨华, 刘黎明, 2017. 297例乳母膳食对乳汁成分的影响[J]. 中国妇幼健康研究, 28(8): 897-899.
|
| [23] |
叶子晴, 赵祥弹, 蔡冰娜, 等, 2022. 牡蛎酶解物减轻5-FU对荷瘤小鼠肠黏膜损伤的作用研究[J/OL]. 热带海洋学报, 41(5): 105-116.
|
| [24] |
曾新斌, 2011. 牡蛎粗多糖对热应激雄性动物血液生化指标及生精功能的影响[D]. 福建: 福建农林大学.
|
| [25] |
张佩, 陈忠琴, 曹文红, 等, 2022. 牡蛎抗氧化及降糖肽的结构特征及其体外模拟消化特性[J]. 食品与发酵工业, 48(21): 196-203.
|
| [26] |
中国发展研究基金会, 2019. 中国母乳喂养影响因素调查报告[R/OL]. (2019-2-28)[2019-5-31]. https://cdrf.org.cn/jjh/pdf/mu.pdf.
CHINA DEVELOPMENT RESEARCH FOUNDATION, 2019. A survey of factors influencing breastfeeding in China[R/OL]. (2019-2-28)[2019-5-31]. https://cdrf.org.cn/jjh/pdf/mu.pdf. (in Chinese with English abstract)
|
| [27] |
朱国萍, 章超桦, 曹文红, 等, 2021. 牡蛎酶解产物对小鼠学习记忆的影响[J]. 广东海洋大学学报, 41(4): 84-92.
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}