
Journal of Tropical Oceanography >
Identification and functional study of the genomic island GIPspSM9913 in Pseudoalteromonas sp. SM9913
Copy editor: YIN Bo
Received date: 2022-05-06
Revised date: 2022-06-29
Online published: 2022-07-01
Supported by
Hainan Provincial Joint Project of Sanya Yazhou Bay Science and Technology City(320LH047)
Natural Science Foundation of Guangdong Province(2019A1515011912)
Science and Technology Planning Project of Guangzhou(202002030493)
Due to the complexity and variability of marine environments, marine bacteria may have evolved unique environmental adaptation mechanisms. Genomic islands usually carry genes related to the environmental adaptation of host bacteria and play an important role in driving bacterial adaptation and genome diversification. Pseudoalteromonas is an important genus that is widely distributed in various marine habitats, and has attracted attention due to the important industrial application and ecological restoration potential. In this study, we focused on Pseudoalteromonas sp. SM9913 isolated from marine sediments. By comparative genomics analysis of SM9913 with the closely related strain Pseudoalteromonas haloplanktis TAC125 isolated from surface seawater, an 18-kb genomic island GIPspSM9913 integrated in the yicC gene was identified. This genomic island encodes an integrase and an excisionase, as well as multiple restriction modification systems. Sequence analysis showed that GIPspSM9913 homologs are widely distributed in marine bacteria such as Pseudoalteromonas, Shewanella, Vibrio and Aeromonas. Quantitative PCR assays showed that GIPspSM9913 can excise when the excisionase is produced, resulting in the removal of GIPspSM9913. Sequencing analysis indicated that excision of GIPspSM9913 did not affect the expression of flanking genes. We also compared the swimming motility, electroporation efficiency and conjugation efficiency of the GIPspSM9913 deleted strain and the wild-type strain SM9913, and found that the presence of GIPspSM9913 can increase the swimming motility of the host bacteria and offer the host defense against the invasion of foreign DNA.
WANG Pengxia , ZHAO Yi , DU Xiaofei , WANG Weiquan , WANG Xiaoxue . Identification and functional study of the genomic island GIPspSM9913 in Pseudoalteromonas sp. SM9913[J]. Journal of Tropical Oceanography, 2023 , 42(2) : 45 -53 . DOI: 10.11978/2022101
表1 本研究所用的引物Tab. 1 The primers used in this study |
| 引物 | 序列 (5′—3′) | 用途 |
|---|---|---|
| xis-F | CGGAATTCATGAACAAACTAATTAAACCAAC | pXis的构建和ΔGIPspSM9913的验证 |
| xis-R | CCCAAGCTTTCATAAAGTGGAGCTCATG | |
| GI-wF | GCCTGGTGTTATGGAAGCAC | ΔGIPspSM9913的验证 |
| GI-wR | GATATTGATTGGCAAGGCGC | |
| GI-qF | GCTGTAGAGCTAAAAGTCCTGATTGA | 定量(附着位点B, attachment site B) attB, 计算GIPspSM9913的切离率 |
| GI-qR | CCAGAAGGCGCAGATAAAATAAATA | |
| gyrB-qF | AGTGGCAACGCTAATTACTGCAT | 定量SM9913基因组上内参基因gyrB, 计算 GIPspSM9913的切离率 |
| gyrB-qR | CGGTAGAAGAACGTTAGTAGCAAGGT |
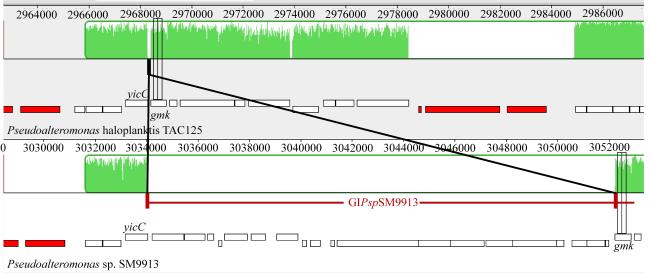
图1 SM9913中基因岛GIPspSM9913的鉴定利用Mauve软件对SM9913和TAC125进行比较基因组学分析, 定位基因岛GIPspSM9913 (Darling et al, 2004) Fig. 1 Identification of genomic island GIPspSM9913 in SM9913. GIPspSM9913 was identified by comparing the genome sequence of SM9913 with TAC125 using Mauve |
表2 基因岛GIPspSM9913上所含基因的信息Tab. 2 The genes in genomic island GIPspSM9913 |
| 基因名称 | 基因编号 | 产物长度/氨基酸数 | 最相似蛋白[菌株](蛋白编号) | 一致性/% | 功能/结构域 |
|---|---|---|---|---|---|
| int | PSM_A2839 | 409 | 整合酶[Pseudoalteromonas atlantica] (WP_024601833.1) | 99 | P4家族整合酶 |
| cds1 | PSM_A2840 | 264 | 假定蛋白[Pseudoalteromonas spiralis] (KYL33016.1) | 99 | 假定蛋白 |
| Xis | PSM_A2841 | 77 | 转录调控因子[Pseudoalteromonas spiralis] (KYL33017.1) | 99 | 切离酶, 原噬菌体CP4-57的调控蛋白(AlpA) |
| cds2 | PSM_A2842 | 38 | - | - | 假定蛋白 |
| cds3 | PSM_A2843 | 293 | 假定蛋白[Pseudoalteromonas spiralis] (KYL33019.1) | 88 | 未知功能结构域(DUF3296) |
| cds4 | PSM_A2844 | 188 | 假定蛋白[Pseudoalteromonas spiralis] (KYL33020.1) | 98 | 假定蛋白 |
| cds5 | PSM_A2845 | 276 | 假定蛋白[Pseudoalteromonas] (WP_024601828.1) | 99 | LabA_like蛋白 |
| cds6 | PSM_A2846 | 56 | 假定蛋白[Paraglaciecola chathamensis S18K6] (GAC12573.1) | 55 | 双功能抗毒素、转录抑制因子RelB |
| cds7 | PSM_A2847 | 126 | 假定蛋白[Pseudoalteromonas] (WP_024601827.1) | 100 | 假定蛋白 |
| cds8 | PSM_A2848 | 44 | HsdR家族Ⅰ型限制修饰系统中的脱氧核糖核酸酶, 部分序列[Pseudoalteromonas arctica A 37-1-2] (ERG10608.1) | 95 | HsdR家族Ⅰ型限制修饰系统中的脱氧核糖核酸酶 |
| hsdR | PSM_A2849 | 1057 | DEAD/DEAH家族解旋酶[Marinospirillum minutulum] (WP_027849446.1) | 78 | HsdR家族Ⅰ型限制修饰系统中的脱氧核糖核酸酶 |
| cds9 | PSM_A2850 | 412 | SIR2家族蛋白[Pseudogulbenkiania sp. MAI-1] (WP_024304176.1) | 43 | SIR2-like结构域蛋白 |
| hsdS | PSM_A2851 | 437 | 限制性内切酶S亚基[Shewanella sp. Sh95] (WP_055647049.1) | 59 | Ⅰ型限制修饰系统中HsdS蛋白,EcoKI限制性修饰系统蛋白 |
| cds10 | PSM_A2852 | 355 | 50S核糖体蛋白L31 [Pseudoalteromonas haloplanktis] (WP_050582305.1) | 95 | 限制性内切酶类(RecB)超家族的核酸酶 |
| hsdM | PSM_A2853 | 559 | DNA甲基化酶[Thalassolituus oleivorans R6-15] (AHK15533.1) | 77 | Ⅰ型限制性修饰系统中N-6 DNA甲基化酶 |
| Mrr | PSM_A2854 | 92 | 限制性内切酶[Pseudoalteromonas sp. S2292] (KJZ24913.1) | 96 | 含有(D/E)-(D/E)XK 活性位点的Ⅱ型限制酶,限制性内切酶 Mrr |
| cds11 | PSM_A2855 | 195 | 假定蛋白[Pseudoalteromonas] (WP_052201246.1) | 95 | 假定蛋白 |
| cds12 | PSM_A2856 | 234 | 大蛋白[Pseudoalteromonas spiralis] (KYL33168.1) | 97 | 具有保守 AHH的HNH/ENDO Ⅶ位点的核酸酶 |
| cds13 | PSM_A2857 | 38 | - | - | 假定蛋白 |
注: -表示 在NCBI中进行BlastP检索未找到相似性蛋白, 无一致性 |
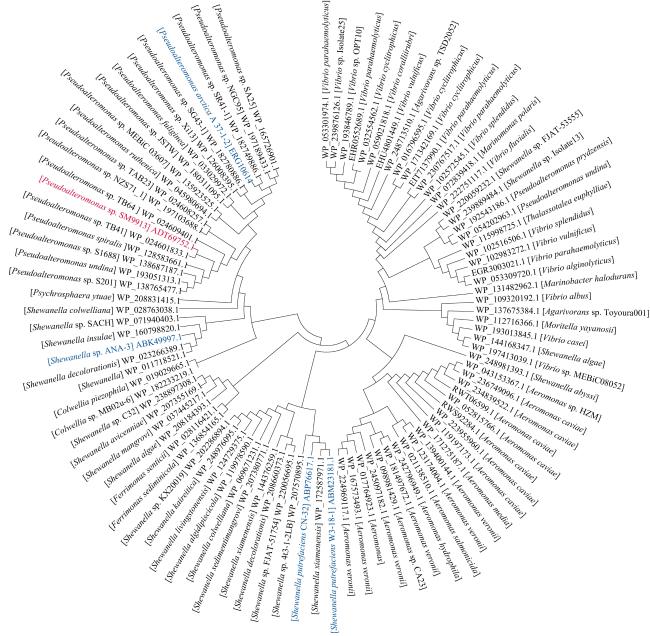
图2 利用MEGA4x1软件采用邻接法对GIPspSM9913编码的整合酶和97个同源蛋白构建系统发育树图中蓝色字体标出的是 Fig. 2 Phylogenetic analysis using the neighbor-joining method to compare the amino acid sequences of the integrase of GIPspSM9913 with 97 homologs. The tree was generated by MEGA4x1. The genomic island analyzed in |
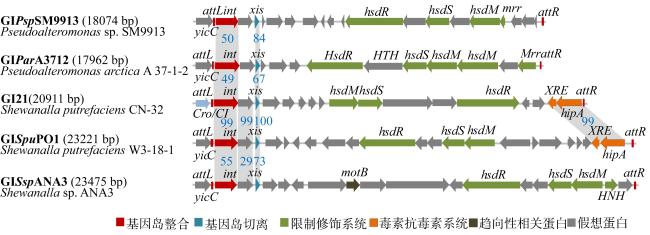
图3 GIPspSM9913 (CP001796.1)与来自假交替单胞菌A 37-1-2 (AHBY00000000.2)、希瓦氏菌CN32 (CP000681.1)、W3-18-1 (CP000503.1)和ANA3 (CP000469.1)的近缘基因岛的多序列比对图中的数字表示同源蛋白之间的序列一致性 Fig. 3 Comparisons of GIPspSM9913 and its related genomic islands. GIPspSM9913 (CP001796.1) was compared with the related genomic islands in Pseudoalteromonas arctica 37-1-2 (AHBY00000000.2), Shewanella putrefaciens CN32 (CP000681), Shewanella putrefaciens W3-18-1 (CP000503) and Shewanella sp. ANA3 (CP000469.1). The percentage of identification between orthologous proteins were shown in the middle. Predicted open reading frames with putative functions are shown in different colors: red, GI recombination; blue, GI excision; green, restriction-modification system; orange, putative toxin-antitoxin system; brown, chemotaxis-related proteins; grey, hypothetical protein |
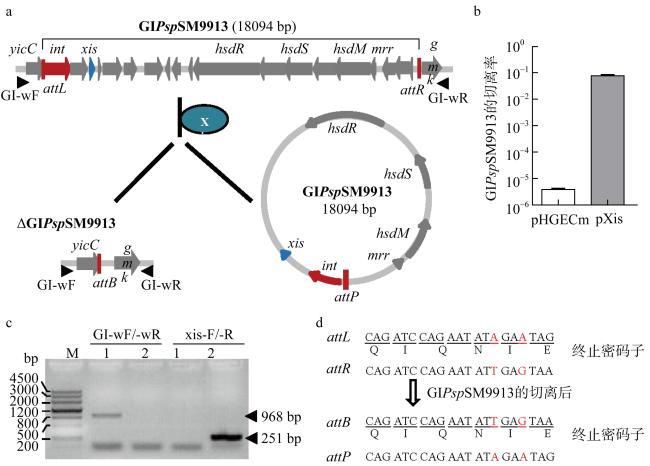
图4 基因岛GIPspSM9913可从SM9913基因组发生切离a. GIPspSM9913从细菌基因组上切离的示意图; b. 切离酶表达时GIPspSM9913的切离率; c. GIPspSM9913切离突变株的PCR验证, DNA模板: 1为突变株ΔGIPspSM9913, 2为野生型SM9913; d. 切离前后GIPspSM9913附着位点的序列比较, 其中红色部分表示序列不一致的碱基 Fig. 4 GIPspSM9913 can excise from the SM9913 genome. (a) schematic of the excision of GIPspSM9913; (b) the excision rate of GIPspSM9913 when the excisionase was overexpressed; (c) PCR confirmation of GIPspSM9913 deletion mutant; the DNA template is: 1, ΔGIPspSM9913, 2, wild-type SM9913; (d) sequence comparison of the attachment sites of GIPspSM9913 before and after its excision |
图5 GIPspSM9913切离对宿主菌生理功能的影响比较野生型菌株(SM9913)和基因岛切离突变株ΔGIPspSM9913的泳动能力(a)、接合转移效率(b)和电转化效率(c) Fig. 5 Effects of GIPspSM9913 excision on the physiological function of host bacteria. (a) the swimming mobility; (b) conjugation efficiency; (c) electroporation efficiency of ΔGIPspSM9913 compared its wild-type strain SM9913 |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}