
Journal of Tropical Oceanography >
Leaves, stems and roots stoichiometry characteristics of mangrove plants at different succession stages in the Shankou National Mangrove Nature Reserve, China*
Copy editor: LIN Qiang
Received date: 2022-05-23
Revised date: 2022-07-28
Online published: 2022-08-02
Supported by
National Natural Science Foundation of China(42049911)
National Natural Science Foundation of China(42141016)
Scientific Research Fund of the Fourth Institute of Oceanography, MNR(202005)
Research Fund Program of Guangxi Key Lab of Mangrove Conservation and Utilization(GKLMC-17A01)
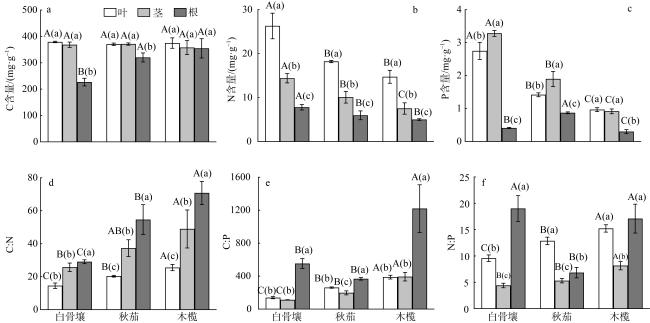
Mangrove is one of the typical coastal wetland ecosystems. Studying its ecological stoichiometry characteristics is important to understand the nutrient utilization, distribution and environmental adaptation strategies of mangrove plants at different successional stages. In the present study, three typical mangrove species, Avicennia marina, Kandelia obovate, and Bruguiera gymnorrhiza were studied. The stoichiometry characteristics of carbon (C), nitrogen (N), and phosphorus (P) in different organs and the sedimental organic carbon (SOC), total nitrogen (TN), and total phosphorus (TP) in the sediments were measured and compared. The ecological stoichiometry characteristics of different organs and sediment nutrients in three intertidal zones of mangrove plants were also analyzed. The results showed that the C, N, and P contents of the three mangrove plants were higher in the leaves and stems than those in the roots(P<0.05). The N and P contents in the leaves and stems of A. marina were significantly higher than those of K. obovate and B. gymnorrhiza (P<0.05). Mangrove plants have unique adaptation strategies to habitats in the different intertidal zones. At the early stage of succession, the plants were distributed in low tide with a competitive strategy. At the advanced stage of succession, the plants were distributed in high tide and the strategy was changed to defense. Correlation analysis showed that the P content in the stems and roots of K. obovate was negatively correlated (P<0.05), while the N and P contents in the leaves and stems were positively correlated (P<0.05). The contents of N and P in the leaves and stems were negatively correlated with SOC, TP and C:N. It was also found that with the succession of mangrove communities from the sea towards the land, the limiting element for the mangrove plants growth has changed from N to both N and P.
Key words: mangrove plant; succession; adaptation strategy; ecological stoichiometry
QIU Jin , DAI Hongtao , XING Yongze , HUANG Daji , Yin Qunjian , CHENG Dewei . Leaves, stems and roots stoichiometry characteristics of mangrove plants at different succession stages in the Shankou National Mangrove Nature Reserve, China*[J]. Journal of Tropical Oceanography, 2023 , 42(3) : 149 -157 . DOI: 10.11978/2022119
表1 样方点经纬度Tab. 1 Latitude and longitude of the sampling sites |
| 主要树种 | 样方号 | 纬度 | 经度 |
|---|---|---|---|
| 白骨壤 | 1 | 21°29′26.257''N | 109°45′59.299''E |
| 2 | 21°29′22.898''N | 109°46′04.188''E | |
| 3 | 21°29′20.515''N | 109°46′07.309''E | |
| 秋茄 | 1 | 21°29′39.548''N | 109°45′50.677''E |
| 2 | 21°29′33.562''N | 109°45′53.338''E | |
| 3 | 21°29′29.541''N | 109°45′54.554''E | |
| 木榄 | 1 | 21°29′53.596''N | 109°45′37.242''E |
| 2 | 21°29′52.703''N | 109°45′37.487''E | |
| 3 | 21°29′51.371''N | 109°45′42.156''E |
表2 山口保护区内三种红树植物概况Tab. 2 Information of three mangrove plants in the Shankou National Mangrove Nature Reserve |
| 植物群落 | 生活型 | 平均基径/cm | 平均胸径/cm | 平均树高/cm | 平均冠幅/cm | 沉积物类型 | 分布区域 |
|---|---|---|---|---|---|---|---|
| 白骨壤 | 灌木或小乔木 | 7.31 | 4.87 | 215 | 258×225 | 淤泥质 | 低潮线 |
| 秋茄 | 小乔木 | 13.36 | 9.45 | 393 | 320×241 | 砂质 | 中潮线 |
| 木榄 | 小乔木 | 23.24 | 11.52 | 378 | 431×351 | 砂质 | 高潮线 |
表3 不同演替阶段沉积物的生态化学计量特征Tab. 3 Ecological stoichiometry characteristics of soil at the different succession stages |
| 植物群落 | SOC/(g·kg-1) | TN/(g·kg-1) | TP/(g·kg-1) | C:N | C:P | N:P |
|---|---|---|---|---|---|---|
| 白骨壤 | 12.85±3.14b | 0.66±0.18b | 0.35±0.11b | 19.45±1.21b | 38.58±10.93a | 1.97±0.44a |
| 秋茄 | 40.11±11.78a | 1.61±0.26a | 0.61±0.06a | 24.48±4.07ab | 67.00±22.39a | 2.68±0.50a |
| 木榄 | 34.46±11.91a | 1.27±0.44ab | 0.53±0.08a | 27.00±1.54a | 68.28±34.30a | 2.53±1.28a |
注: 表中数据为平均值±标准差, 上标不同字母表示不同植物群落间差异显著, 字母顺序表示均值大小关系, 下同。设置显著性水平为P<0.05 |
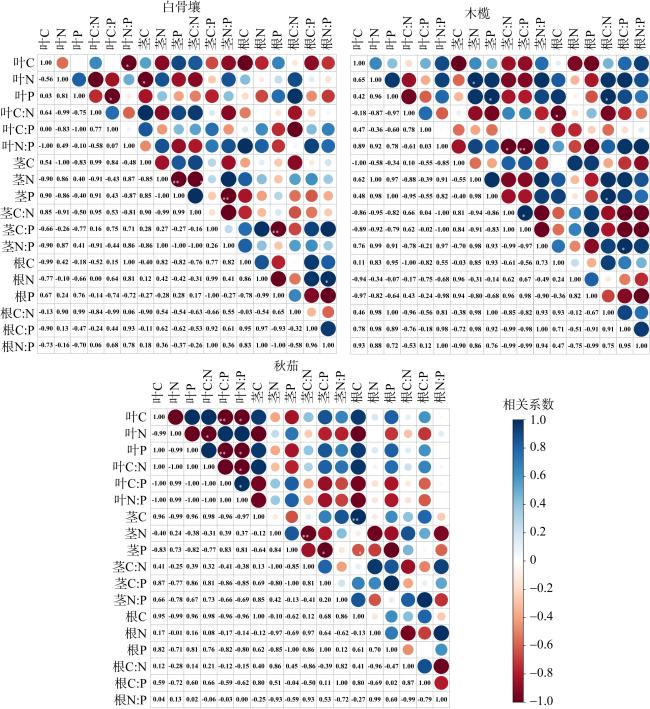
图2 不同演替阶段三种红树植物叶、茎、根C、N、P及计量比相关性热图图中右上部分为相关性图, 蓝色表示正相关关系, 红色表示负相关关系, 圆圈的颜色深浅表示相关系数的绝对值大小, 圆圈的大小表示相关性大小。图中左下部分为相关系数数值, *表示相关性显著(P<0.05); **表示相关性极显著(P<0.01) Fig. 2 The correlation heat map of C, N, P content and stoichiometric ratio of the leaves, stems and roots of mangrove plants at different succession stages |
表4 红树植物化学计量比与沉积物养分含量及计量特征的相关性Tab. 4 The correlations of C, N, P ecological stoichiometry of the mangrove plant and soil |
| 植物元素 | 器官 | 沉积物养分含量及计量比 | |||||
|---|---|---|---|---|---|---|---|
| SOC | TN | TP | C:N | C:P | N:P | ||
| C | 叶 | -0.26 | -0.25 | -0.40 | -0.26 | -0.07 | 0.00 |
| 茎 | 0.11 | 0.13 | 0.18 | -0.02 | -0.03 | -0.04 | |
| 根 | 0.83** | 0.76* | 0.53 | 0.83** | 0.76* | 0.61 | |
| 植物元素 | 器官 | 沉积物养分含量及计量比 | |||||
| SOC | TN | TP | C:N | C:P | N:P | ||
| N | 叶 | -0.69* | -0.63 | -0.81** | -0.78* | -0.42 | -0.21 |
| 茎 | -0.68* | -0.60 | -0.75* | -0.85** | -0.44 | -0.22 | |
| 根 | -0.54 | -0.50 | -0.54 | -0.67* | -0.45 | -0.31 | |
| P | 叶 | -0.71* | -0.67* | -0.78* | -0.77* | -0.48 | -0.28 |
| 茎 | -0.69* | -0.60 | -0.61 | -0.88** | -0.55 | -0.34 | |
| 根 | 0.44 | 0.54 | 0.51 | 0.05 | 0.17 | 0.20 | |
| C:N | 叶 | 0.58 | 0.49 | 0.68* | 0.80** | 0.37 | 0.14 |
| 茎 | 0.54 | 0.42 | 0.66 | 0.82** | 0.30 | 0.06 | |
| 根 | 0.67* | 0.61 | 0.51 | 0.76* | 0.61 | 0.46 | |
| C:P | 叶 | 0.56 | 0.48 | 0.60 | 0.77* | 0.40 | 0.20 |
| 茎 | 0.49 | 0.35 | 0.45 | 0.82** | 0.39 | 0.17 | |
| 根 | 0.15 | 0.04 | -0.08 | 0.42 | 0.32 | 0.23 | |
| N:P | 叶 | 0.61 | 0.55 | 0.57 | 0.71* | 0.49 | 0.33 |
| 茎 | 0.46 | 0.35 | 0.23 | 0.71* | 0.52 | 0.36 | |
| 根 | -0.58 | -0.65 | -0.67* | -0.31 | -0.29 | -0.26 | |
注: 加粗字段表示存在显著相关性, *代表相关性显著(P<0.05), **代表相关性极显著(P<0.01) |
表5 不同研究区域植物叶片含量及生态化学计量特征比较Tab. 5 Comparison of leaf element contents and ratios in the different study areas |
| 研究区域 | C含量/(mg·g-1) | N含量/(mg·g-1) | P含量/(mg·g-1) | C:N | C:P | N:P | 数据来源 |
|---|---|---|---|---|---|---|---|
| 全球植物 | — | 20.62 | 1.99 | 22.50 | 232.00 | 12.70 | Elser等(2000) |
| 全球陆生植物 | 476.10 | 17.40 | 1.23 | 23.40 | — | 12.80 | Kattge等(2011) |
| 中国陆生植物 | — | 20.24 | 1.46 | — | — | 16.30 | Han等(2005) |
| 中国陆生植物 | — | 19.09 | 1.56 | — | — | 15.39 | 任书杰 等(2007) |
| 中国湿地 | — | 18.30 | 2.55 | — | — | 7.18 | Hu等(2021) |
| 中国东部湿地 | 369.00 | 25.90 | 3.28 | 17.10 | 149.00 | 9.50 | Xia等(2014) |
| 中国湿地植物 | — | 16.07 | 1.85 | — | — | 8.67 | 胡伟芳 等(2014) |
| 全球滨海湿地 | 418.00 | 16.13 | 1.59 | 37.09 | 432.20 | 13.04 | Hu等(2021) |
| 广西山口 | 374.83 | 19.75 | 1.71 | 20.15 | 262.88 | 12.55 | 本研究 |
| [1] |
陈亮, 洪文君, 黄永平, 等, 2019. 6种红树树种生长与叶片元素含量及化学计量特征[J]. 林业与环境科学, 35(1): 83-88.
|
| [2] |
樊月, 潘云龙, 陈志为, 等, 2019. 四种红树植物根茎叶的碳氮磷化学计量特征[J]. 生态学杂志, 38(4): 1041-1048.
|
| [3] |
郭子武, 陈双林, 杨清平, 等, 2012. 雷竹林土壤和叶片N、P化学计量特征对林地覆盖的响应[J]. 生态学报, 32(20): 6361-6368.
|
| [4] |
何斌, 温远光, 梁宏温, 等, 2002. 英罗港红树植物群落不同演替阶段植物元素分布及其与土壤肥力的关系[J]. 植物生态学报, 26(5): 518-524.
|
| [5] |
何琴飞, 申文辉, 彭玉华, 等, 2017. 钦州湾红树林土壤肥力及其C、N、P、K化学计量特征[J]. 西北林学院学报, 32(6): 119-124, 149.
|
| [6] |
胡伟芳, 章文龙, 张林海, 等, 2014. 中国主要湿地植被氮和磷生态化学计量学特征[J]. 植物生态学报, 38(10): 1041-1052.
|
| [7] |
黄建军, 王希华, 2003. 浙江天童32种常绿阔叶树叶片的营养及结构特征[J]. 华东师范大学学报(自然科学版), 2003(1): 92-97.
|
| [8] |
黎洁, 2017. 广西北仑河口红树林植物种群动态与生态化学计量特征研究[D]. 广西师范学院.
|
| [9] |
卢妍, 2010. 湿地植物对淹水条件的响应机制[J]. 自然灾害学报, 19(4): 147-151.
|
| [10] |
罗艳, 贡璐, 2016. 塔里木盆地南缘不同生境下芦苇生态化学计量特征[J]. 生态学杂志, 35(3): 684-691.
|
| [11] |
邱岭军, 何宗明, 胡欢甜, 等, 2017. 滨海沙地不同树种碳氮磷化学计量特征[J]. 应用与环境生物学报, 27(3): 555-559.
|
| [12] |
任书杰, 于贵瑞, 陶波, 等, 2007. 中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究[J]. 环境科学, 28(12): 2665-2673.
|
| [13] |
宋同清, 彭晚霞, 杜虎, 等, 2014. 中国西南喀斯特石漠化时空演变特征、发生机制与调控对策[J]. 生态学报, 34(18): 5328-5341.
|
| [14] |
苏柏予, 张维仕, 王友绍, 2021. 三种红树根部组织抗氧化酶活性对水淹胁迫的响应[J]. 热带海洋学报, 41(6): 35-43.
|
| [15] |
魏晨, 张小平, 罗子渝, 等, 2021. 兰州市南山和北山3种乔木叶片生态化学计量特征的对比研究[J]. 生态学报, 41(6): 2460-2470.
|
| [16] |
余泺, 高明, 王子芳, 等, 2011. 土壤水分对烤烟生长、物质分配和养分吸收的影响[J]. 植物营养与肥料学报, 17(4): 989-995.
|
| [17] |
张庆费, 宋永昌, 由文辉, 1999. 浙江天童植物群落次生演替与土壤肥力的关系[J]. 生态学报, 19(2): 174-178.
|
| [18] |
周丽丽, 李树斌, 王万萍, 等, 2020. 福建漳江口4种红树植物叶片碳氮磷化学计量及养分重吸收特征[J]. 应用与环境生物学报, 26(3): 674-680.
|
| [19] |
周元慧, 张忠华, 黎洁, 等, 2020. 广西北仑河口红树植物叶片和土壤的碳氮磷生态化学计量特征[J]. 地球与环境, 48(1): 58-65.
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}