
Journal of Tropical Oceanography >
Interactions among Aureococcus anophagefferens, Skeletonema costatum, and Chattonella marina under different temperatures
Copy editor: LIN Qiang
Received date: 2020-12-29
Request revised date: 2021-02-09
Online published: 2021-02-10
Supported by
National Natural Science Foundation of China(41876173)
Foundation of Department of Natural Resources of Heibei Province(454-0503-JBN-XCLS)
Copyright
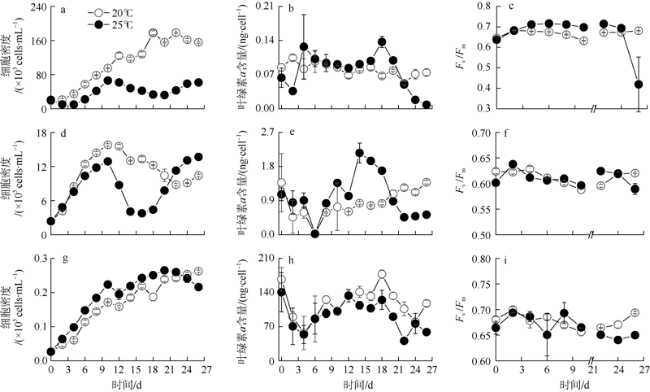
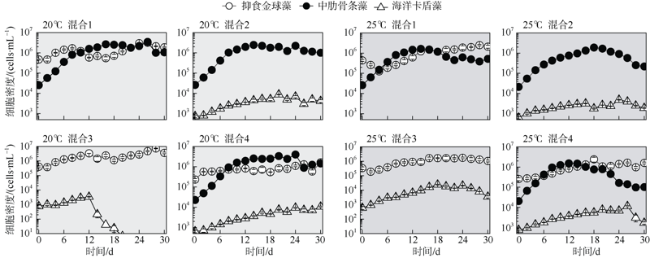
Mono-, bi-, and tri-algal cultures were applied to Aureococcus anophagefferens, Skeletonema costatum, and Chattonella marina under two temperatures of 20℃ and 25℃, to investigate species-specific interaction and competition in different temperatures. Single cultures showed the environmental carry capacity (K) for A. anophagefferens and S. costatum were significantly reduced by increased temperature, indicating they are sensitive to rising temperature. In contrast, the inherent growth rate (r) for C. marina significantly increased at 25℃ whereas K remained the same as that at 20℃. In contrast to the death under A. anophagefferens addition at 20℃, K of C. marina increased 120% at 25℃. Although r of S. costatum was promoted by C. marina addition at both temperatures, its K showed from 43% increase to 48% decrease from 20℃ to 25℃. The change of temperature did not influence the results of co-culture of S. costatum and A. anophagefferens with 40% increases and 60%-70% decreases of K at both temperatures. The results of bi-cultures indicated that interspecies competition is not only influenced by fixed interaction relationship of co-cultured species, but also by difference of algal temperature adaptability. In tri-algal cultures at 20℃, the inhibition rate of A. anophagefferens and the stimulation rate of S. costatum both reached the highest of 79% and 108%, respectively, which probably illuminated a synergistic effect within three algal interaction.
ZHENG Chengzhi , ZUO Liming , MA Wang , ZHU Qin , WANG Huohuo , LÜ Songhui , CHEN Heng , HUANG Kaixuan . Interactions among Aureococcus anophagefferens, Skeletonema costatum, and Chattonella marina under different temperatures[J]. Journal of Tropical Oceanography, 2021 , 40(3) : 124 -131 . DOI: 10.11978/2020154
表1 不同混合处理的初始密度和混合比例Tab. 1 Initial cell intensities and mixture ratios of various combination cultures |
| 处理 | 初始密度/(cells·mL-1) | 混合比例 | ||
|---|---|---|---|---|
| 抑食金球藻 | 中肋骨条藻 | 海洋卡盾藻 | ||
| 单种 | 20×105 | 2.5×105 | 2.5×103 | N/A |
| 混合1 | 40×105 | 2.5×105 | N/A | 16∶1 |
| 混合2 | N/A | 2.5×105 | 750 | 33∶1 |
| 混合3 | 40×105 | N/A | 750 | 530∶1 |
| 混合4 | 25×105 | 2.5×105 | 750 | 333∶33∶1 |
注: N/A表示细胞未参与共培养 |
表2 不同温度和混合处理下三种藻的生长参数Tab. 2 Parameters of growth for three algae species under various temperature and combination treatments |
| 温度/℃ | 种名 | 处理 | 环境容纳量K/(×105 cells·mL-1) | 內禀增长率r | 生长拐点Tp/d |
|---|---|---|---|---|---|
| 20 | 抑食金球藻 | 单种 | 161.63±3.40 | 0.24±0.02 | 8.54±0.63 |
| 混合1 | 48.09±15.50 | 0.29±0.07 | 23.83±4.76 | ||
| 混合3 | 83.31±71.28 | 0.24±0.07 | 13.46±3.34 | ||
| 混合4 | 34.72±4.80 | 0.43±0.01 | 46.00±0.85 | ||
| 海洋卡盾藻 | 单种 | 0.25±0.01 | 0.26±0.02 | 7.77±0.68 | |
| 混合2 | 0.09±0.02 | 0.20±0.02 | 13.2±1.11 | ||
| 混合3 | 0.07±0.04 | 0.23±0.06 | 49.00±7.67 | ||
| 混合4 | 0.10±0.02 | 0.14±0.02 | 18.50±1.32 | ||
| 中肋骨条藻 | 单种 | 16.66±0.53 | 0.45±0.02 | 4.04±0.15 | |
| 混合1 | 23.28±3.04 | 0.45±0.03 | 10.20±0.46 | ||
| 混合2 | 23.84±1.99 | 0.52±0.02 | 9.12±0.25 | ||
| 混合4 | 34.73±4.79 | 0.43±0.01 | 11.77±0.3 | ||
| 25 | 抑食金球藻 | 单种 | 65.21±4.93 | 0.63±0.17 | 6.89±1.78 |
| 混合1 | 27.10±3.57 | 0.19±0.02 | 19.42±1.34 | ||
| 混合3 | 19.57±2.68 | 0.24±0.03 | 11.42±1.03 | ||
| 混合4 | 18.57±6.40 | 0.17±0.04 | 13.00±2.44 | ||
| 海洋卡盾藻 | 单种 | 0.25±0.01 | 0.40±0.03 | 5.10±0.35 | |
| 混合2 | 0.05±0.01 | 0.15±0.02 | 10.07±1.2 | ||
| 混合3 | 0.55±0.50 | 0.20±0.02 | 21.85±3.32 | ||
| 混合4 | 0.13±0.03 | 0.15±0.02 | 18.00±1.73 | ||
| 中肋骨条藻 | 单种 | 13.37±0.33 | 0.47±0.01 | 3.23±0.07 | |
| 混合1 | 18.70±2.56 | 0.49±0.02 | 8.88±0.27 | ||
| 混合2 | 6.99±0.39 | 0.52±0.02 | 6.75±0.18 | ||
| 混合4 | 14.94±1.22 | 0.63±0.10 | 7.98±0.64 |
| [1] |
毕相东, 张兴华, 宋伦, 等, 2016. 小球藻与褐潮原因种-抑食金球藻间的相互作用研究[J]. 海洋与湖沼, 47(3):594-603.
|
| [2] |
陈杨航, 梁君荣, 陈长平, 等, 2015. 褐潮——一种新型生态系统破坏性藻华[J]. 生态学杂志, 34(1):274-281.
|
| [3] |
黄娟, 杨维东, 刘洁生, 等, 2009. 温度、盐度、光照对海洋卡盾藻生长和产毒的影响[J]. 应用生态学报, 20(5):1190-1195.
|
| [4] |
雷蕾, 姚鹏, 2016. 抑食金球藻褐潮环境影响因素研究进展[J]. 海洋环境科学, 35(4):635-640.
|
| [5] |
刘望鹏, 黄娟, 杨维东, 等, 2010. 营养盐对海洋卡盾藻生长与产毒的影响[J]. 生态毒理学报, 5(3):394-401.
|
| [6] |
吕颂辉, 李英, 2006. 我国东海4种赤潮藻的细胞氮磷营养储存能力对比[J]. 过程工程学报, 6(3):439-444.
|
| [7] |
潘克厚, 王金凤, 朱葆华, 2007. 海洋微藻间竞争研究进展[J]. 海洋科学, 31(5):58-62.
|
| [8] |
王朝晖, 袁美玲, 骆育敏, 等, 2010. 海洋卡盾藻与中肋骨条藻和锥状斯氏藻种间竞争研究[J]. 水生生物学报, 34(2):336-344.
|
| [9] |
张达娟, 张兴华, 唐学玺, 等, 2017. 三角褐指藻与抑食金球藻的竞争及化感作用研究[J]. 海洋学报, 39(6):84-94.
|
| [10] |
张磊, 李航霄, 吴敏, 等, 2020. 不同温度下海水酸化对中肋骨条藻光合生理特性的影响[J]. 江苏海洋大学学报(自然科学版), 29(1):1-7.
|
| [11] |
张小华, 刘东艳, 2020. 温度与溶解态有机磷源对中肋骨条藻生长的影响[J]. 海洋科学, 44(11):36-44.
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}