
Journal of Tropical Oceanography >
Effect of suspended solids on zooplankton community and their feeding selectivity in the Sanya coral waters
Copy editor: LIN Qiang
Received date: 2022-06-09
Revised date: 2022-07-22
Online published: 2022-08-21
Supported by
National Natural Science Foundation of China(41806188)
National Natural Science Foundation of China(42176118)
Science and Technology Planning Project of Guangdong Province(2020B1212060058)
Hainan Provincial Natural Science Foundation of China(422QN442)
Science and Technology Planning Project of Guangzhou City(202201010674)
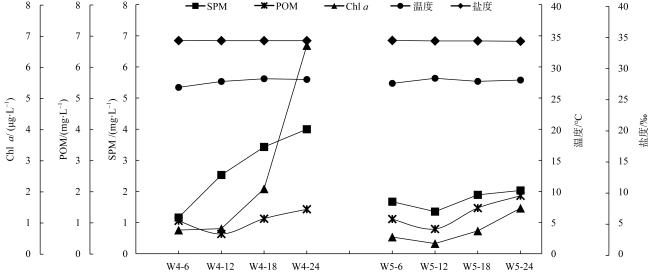
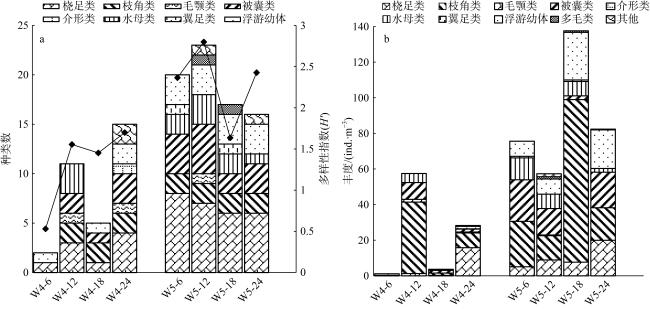
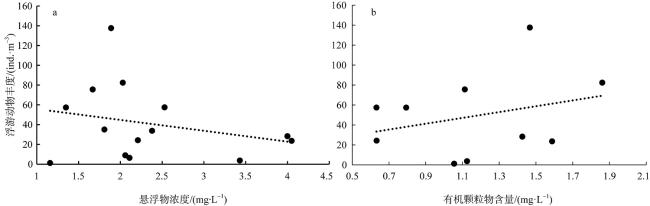
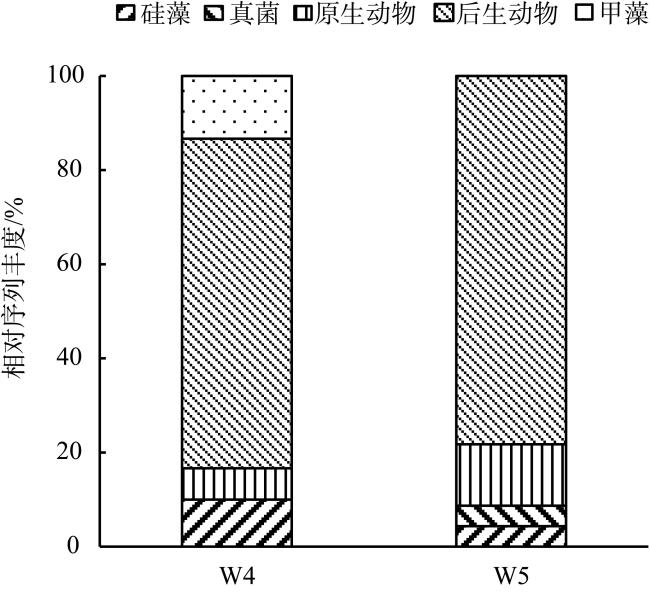
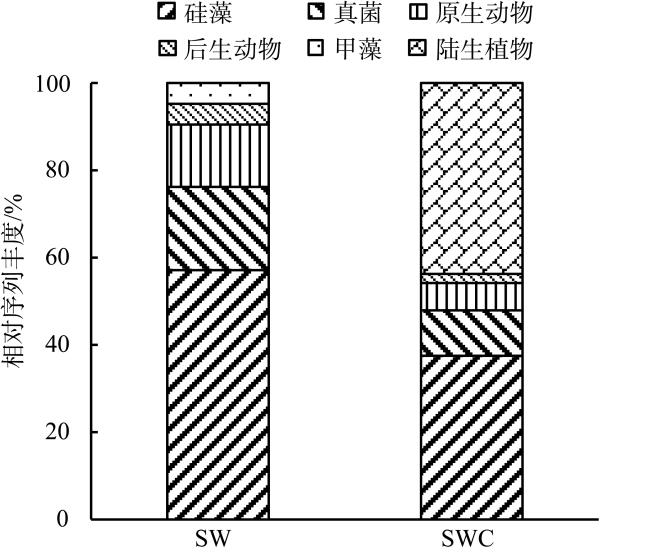
With the intensification of human activities, the suspended solids imported into the nearshore coral reef area are increasing, which will not only affect the physiology of coral but also affect the basic biological production process of the coral reef ecosystem. To understand the impact of suspended solids on the species composition and feeding of zooplankton, the impact of suspended solids fluctuation on zooplankton community structure was analyzed by a diurnal observation in a typical coastal reef area of Sanya in April 2015, and the feeding response of the dominant species was studied by molecular method combined with laboratory experiments. The results showed that the phytoplankton biomass (chlorophyll a concentration) in these two stations was low with a significant diurnal fluctuation. The concentration of suspended solids and chlorophyll a in the W4 station are higher than those of the W5 station, but the proportion of organic particulate matter in the suspended solids of the W5 (73.69%) station is higher than that of W4 (46.09%). The species diversity and abundance of zooplankton in the W5 station are higher than that in the W4 station. Dietary analysis shows that the copepods in these two stations are mainly fed on organic detritus from metazoans. The proportion of phytoplankton in the diet of copepods in the W4 station is also relatively high, while copepods in the W5 station also eat some microzooplankton. In the feeding experiment, the high concentration of suspended particles significantly improved the filtration rate and ingestion rate of Acartia erythraea, but its dominant food item changed from phytoplankton to terrestrial plant detritus, indicating that the high content of suspended particles in the concentrated seawater limited the feeding choice of copepods and made them eat more organic detritus as a compensatory food source. The results show that high suspended solids concentration will affect the feeding choice of zooplankton, but the high proportion of organic detritus in suspended solids can provide an important supplementary food source for zooplankton and support its maintenance of high biomass.
Key words: zooplankton; suspended solid; community structure; diet diversity; Sanya Bay
HU Simin , ZHOU Tiancheng , ZHANG Chen , LIU Sheng , LI Tao , HUANG Hui . Effect of suspended solids on zooplankton community and their feeding selectivity in the Sanya coral waters[J]. Journal of Tropical Oceanography, 2024 , 43(3) : 122 -130 . DOI: 10.11978/2022132
表 1 采样站点相关参数Tab. 1 Information of sampling sites |
| 站位 | 经纬度 | 水深/m | 分布区域 | 调查日期 |
|---|---|---|---|---|
| W4 | 18°13′N, 109°28′E | 7 | 礁区上方 | 04-18 |
| W5 | 18°12′N, 109°25′E | 15 | 礁区边缘 | 04-17 |
表 2 桡足类现场摄食相关指数Tab. 2 Indexs of copepod in situ diets |
| 样品号 | 种类数 | 有效克隆数 | 香农-威纳指数 | Chao1指数a | 杂食指数 | 食谱宽度 |
|---|---|---|---|---|---|---|
| W4 | 13 | 31 | 1.791 | 28 | 0.76 | 3.28 |
| W5 | 10 | 26 | 1.881 | 31 | 0.8 | 5.24 |
注: a Chao1指数计算公式为Chao1=S+ni(n1-1)/2(n2+1), 式中S是得到的种类数量, n1为只有一条序列的序列数目, n2为只有两条序列的序列数目(Chao, 1984) |
表3 不同摄食实验组中红纺锤水蚤的摄食率和滤水率Tab. 3 Grazing rate and filter rate of A. erythraea at different incubation treatments |
| 处理组 | 实验前细胞丰度/(×104cells∙L-1) | 实验后细胞丰度/(×103cells∙L-1) | 桡足类个体数 | 培养时间 | 滤水率/ (mL∙ind.-1∙h-1) | 摄食率/ (cells∙ind.-1∙h-1) |
|---|---|---|---|---|---|---|
| SW | 1.04 | 3.21±0.01 | 50 | 4h | 0.65±0.03 | 3.98±0.01 |
| SWC | 4.16 | 1.56±0.07 | 50 | 4h | 1.64±0.04 | 20.02±0.07 |
注: SW, 现场海水作为饵料; SWC, 现场海水进行浓缩后作为饵料; 实验中用到的桡足类均为成熟雌性个体 |
| [1] |
高亚辉, 林波, 1999. 几种因素对太平洋纺锤水蚤摄食率的影响[J]. 厦门大学学报(自然科学版), 38(5): 751-757.
|
| [2] |
柯志新, 黄良民, 谭烨辉, 等, 2011. 三亚珊瑚礁分布海区浮游生物的群落结构[J]. 生物多样性, 19(6): 696-701.
|
| [3] |
宋星宇, 谭烨辉, 黄良民, 等, 2011. 三亚湾春夏季初级生产力的分布特征及环境影响研究[J]. 海洋环境科学, 30(1): 19-23.
|
| [4] |
尹健强, 张谷贤, 谭烨辉, 等, 2004. 三亚湾浮游动物的种类组成与数量分布[J]. 热带海洋学报, 23(5): 1-9.
|
| [5] |
赵美霞, 余克服, 施棋, 等, 2013. 三亚鹿回头珊瑚礁区沉降物的来源、分布及影响因素[J]. 科学通报, 58(17): 1583-1589.
|
| [6] |
赵卫东, 2000. 南沙珊瑚礁生态系的营养动力学过程[D]. 青岛: 中国科学院海洋研究所: 4-55.
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}