
Journal of Tropical Oceanography >
Effects of polar and lateral flagella on biofilm formation in marine Pseudoalteromonas
Copy editor: LIN Qiang
Received date: 2022-07-03
Revised date: 2022-08-31
Online published: 2022-09-14
Supported by
National Natural Science Foundation of China(31970037)
National Natural Science Foundation of China(91951203)
Guangdong Major Project of Basic and Applied Basic Research(2019B030302004)
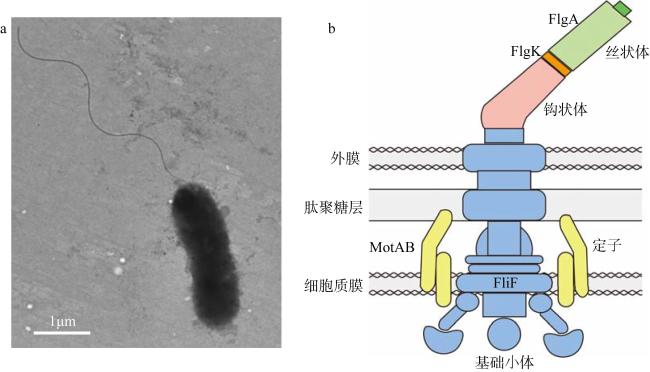
Pseudoalteromonas are widely distributed in diverse marine environments. They can produce an array of bioactive compounds and enzymes, and have strong ability to form biofilms in general. The biofilms formed by Pseudoalteromonas are able to induce biomineralization and inhibit the settlement of fouling organisms. Flagella are essential for bacterial motility, mediating nutrient acquisition, and the transformation from planktonic living style to biofilm state. Flagella is also considered as a critical component of biofilm matrix. However, the function of flagella during biofilm formation of marine Pseudoalteromonas remains unclear. In this study, we focused on Pseudoalteromonas sp. SCSIO 11900 and SM9913, which were isolated from the surface water and the sediment of deep oceans, respectively. The influence of flagella on biofilm formation and motility was explored by analyzing mutants lacking the genes encoding components of different flagellar portions in both strains. The results showed that the polar flagella are essential for swimming motility, and inhibit biofilm formation in SM9913 and SCSIO 11900. In addition, the lateral flagella in SM9913 also plays a role in swimming motility, but has no effects on biofilm formation. We further showed that these polar flagella systems are present in different Pseudoalteromonas strains, implying that they may be important for these bacteria for nutrient acquisition, for colonization and environmental adaptation in diverse marine habitats.
Key words: Pseudoalteromonas; flagella; swimming motility; biofilm formation
GUO Yunxue , CAI Xingsheng , GU Jiayu , WANG Xiaoxue . Effects of polar and lateral flagella on biofilm formation in marine Pseudoalteromonas[J]. Journal of Tropical Oceanography, 2023 , 42(3) : 126 -135 . DOI: 10.11978/2022149
表1 实验中用到的菌株和质粒Tab. 1 Bacterial strains and plasmids used in this study |
| 描述 | 来源 | ||
|---|---|---|---|
| 菌株 | SCSIO 11900 | 野生型, 分离自中国南海4m水深的珊瑚黏液层 | Zeng et al, 2014 |
| 11900ΔmotAB | 敲除motAB的SCSIO 11900菌 | 本研究 | |
| 11900 ΔfliF | 敲除fliF的SCSIO 11900菌 | 本研究 | |
| 11900 ΔflgK | 敲除flgK的SCSIO 11900菌 | 本研究 | |
| 11900 ΔflaA | 敲除flaA的SCSIO 11900菌 | 本研究 | |
| SM9913 | 野生型, 分离自冲绳海槽附近1855m水深的沉积物 | Qin et al, 2011 | |
| 9913ΔmotAB | 敲除motAB的SM9913菌 | 本研究 | |
| 9913 ΔfliF | 敲除fliF的SM9913菌 | 本研究 | |
| 9913 ΔflgK | 敲除flgK的SM9913菌 | 本研究 | |
| 9913 ΔflaA | 敲除flaA的SM9913菌 | 本研究 | |
| ΔLF | 敲除侧生鞭毛基因簇的SM9913菌 | 本研究 | |
| ΔLFΔfliF | 在ΔfliF中敲除侧生鞭毛基因簇基因的SM9913菌 | 本研究 | |
| ΔLFΔflgk | 在ΔflgK中敲除侧生鞭毛基因簇基因的SM9913菌 | 本研究 | |
| ΔLFΔflaA | 在ΔflaA中敲除侧生鞭毛基因簇基因的SM9913菌 | 本研究 | |
| WM3064 | 接合转移用的DAP营养缺陷型大肠杆菌, 37°C | 实验室保存 | |
| 基因敲除质粒 | pK18mobsacB-Ery | pK18mobsacB 质粒插入pHT304来源的红霉素抗性基因, Kanr, Eryr | Wang et al, 2015 |
| pK18Ery-11900 motAB | pK18mobsacB-Ery 携带SCSIO 11900 motAB基因上下游片段 | 本研究 | |
| pK18Ery-11900 fliF | pK18mobsacB-Ery 携带SCSIO 11900 fliF基因上下游片段 | 本研究 | |
| pK18Ery-11900 flgK | pK18mobsacB-Ery 携带SCSIO 11900 flgK基因上下游片段 | 本研究 | |
| pK18Ery-11900 flaA | pK18mobsacB-Ery 携带SCSIO 11900 flaA基因上下游片段 | 本研究 | |
| pK18Ery-9913 motAB | pK18mobsacB-Ery 携带SM9913端生鞭毛 motAB基因上下游片段 | 本研究 | |
| pK18Ery-9913 fliF | pK18mobsacB-Ery 携带SM9913端生鞭毛fliF基因上下游片段 | 本研究 | |
| pK18Ery-9913 flgK | pK18mobsacB-Ery 携带SM9913端生鞭毛flgK基因上下游片段 | 本研究 | |
| pK18Ery-9913 flaA | pK18mobsacB-Ery 携带SM9913端生鞭毛flaA基因上下游片段 | 本研究 | |
| pK18Ery-9913 LF | pK18mobsacB-Ery 携带SM9913侧生鞭毛基因簇上下游片段 | 本研究 |
注: r指抗生素抗性; Kan指卡那霉素; Ery指红霉素 |
表2 实验中用到的引物Tab. 2 Primers used in this study |
| 名称 | 序列 | 用途 |
|---|---|---|
| 11900ΔmotAB-up-F | ACATGCATGCGCTAATGCTGCGAT | 用于敲除SCSIO 11900中的motAB |
| 11900ΔmotAB-up-R | CGGAATTCATTCCCTCCAAAGGTTTTAT | |
| 11900ΔmotAB-down-F | CGGAATTCAATGACTTGGCTATGTACCT | |
| 11900ΔmotAB-down-R | CTAG TCTAGATAAGTCACTGATCAATAAAC | |
| 11900ΔfliF-up-F | ACATGCATGCGCTAATAAAGTTGCTCGATC | 用于敲除SCSIO 11900中的fliF |
| 11900ΔfliF-up-R | CGGAATTCAATAATTCTCCACTACCTAGTT | |
| 11900ΔfliF-down-F | CGGAATTCTCAAGAACAAAAACAACTACC | |
| 11900ΔfliF-down-R | ACGCGTCGACGTACCAGCTGTCTTTCTGCT | |
| 11900ΔflgK-up-F | ACATGCATGCTGGTGGTTTAACAGTCACGA | 用于敲除SCSIO 11900中的flgK |
| 11900ΔflgK-up-R | CGGAATTCGTTACTCTACTCCTTGGCGG | |
| 11900ΔflgK-down-F | CGGAATTCTTATGCGTTTATCAAATAACT | |
| 11900ΔflgK-down-R | ACGCGTCGACCATTGAGGTTCAAACAATAA | |
| 11900ΔflaA-up-F | GACTAACATTGCTTACCGCATGAA | 用于敲除SCSIO 11900中的flaA |
| 11900ΔflaA-up-R | TTTACCATCTCCTAGATTTAAA | |
| 11900ΔflaA-down-F | ACATAGATGTTAGATTTAAATGAT | |
| 11900ΔflaA-down-R | TCCATTAGCAATACTAAATTCATT | |
| 9913ΔmotAB-up-F | ACATGCATGC GCTAGTGCTGCGATCATTT | 用于敲除SM9913中端生鞭毛的motAB |
| 9913ΔmotAB-up-R | CGGAATTCATTCCCTCCAAAGGTTTTA | |
| 9913ΔmotAB-down-F | CGGAATTCAATGACTTGGCTATGTACCT | |
| 9913ΔmotAB-down-R | CTAGTCTAGATTGCTCGATTACTTCATCA | |
| 9913ΔfliF-up-F | AGAAACCTTGGCACGCTGCCA | 用于敲除SM9913中端生鞭毛的fliF |
| 9913ΔfliF-up-R | AATAATTCTCCACTACCT | |
| 9913ΔfliF-down-F | TCAAGAACAAAAACAACTACCTC | |
| 9913ΔfliF-down-R | TCTTCTAAATTAGCAATACGC | |
| 9913ΔflgK-up-F | ACATGCATGC GAATAGTCAATCTGAAGGG | 用于敲除SM9913中端生鞭毛的flgK |
| 9913ΔflgK-up-R | CGGAATTC TTTACCATCTCCTAGTTTCA | |
| 9913ΔflgK-down-F | CGGAATTC TTTAACAATAAGCTCTATACTTT | |
| 9913ΔflgK-down-R | CTAG TCTAGA GATAATACTAACCGCTGCC | |
| 9913ΔflaA-up-F | GTTAGGTGGTTAATAATAATTAAAA | 用于敲除SM9913中端生鞭毛的flaA |
| 9913ΔflaA-up-R | ACGTTGTAGAATGTTTGATGACTATG | |
| 9913ΔflaA-down-F | TTTAACAATAAGCTCTATACTTTA | |
| 9913ΔflaA-down-R | TACCCTCTTAAATTACTGAAAAATT | |
| 9913LF-up-F | ACATGCATGCGCAGTTCGTGCTCTAGGTAA | 用于敲除SM9913中侧生鞭毛基因簇 |
| 9913LF-up-R | CGGAATTC AAAATGTCGAAATAAAAGTT | |
| 9913LF-down-F | CGGAATTC TAAATTTTTAGGATTACAACAA | |
| 9913LF-down-R | ACGCGTCGACCAAGCATCAGTTGGCTAATA |
注: F为正向引物, R为反向引物 |
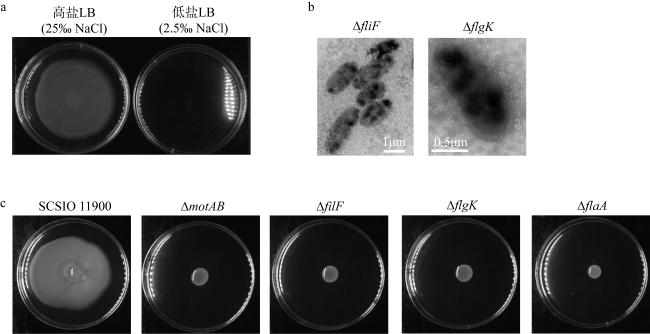
图2 鞭毛基因敲除对细菌游动性的影响a. SCSIO 11900在高盐LB培养基中游动, 在低盐LB培养基中不游动; b. 敲除鞭毛基因fliF和flgK后SCSIO 11900电镜图; c.野生型SCSIO 11900与端生鞭毛基因敲除的菌株在游动性平板上游动24h的情况 Fig. 2 Effects of flagella genes deletion on motility of SCSIO 11900. (a) SCSIO 11900 motile on high salt LB medium other than low salt LB medium; (b) Deletion flagella genes fliF and flgK resulted in the loss of motility of SCSIO 11900 cells; (c) Polar flagella mutant inhibits motility of SCSIO 11900. Wild type SCSIO 11900 and flagella destroyed mutant strains grew in LB medium containing 25‰ NaCl for 24h |
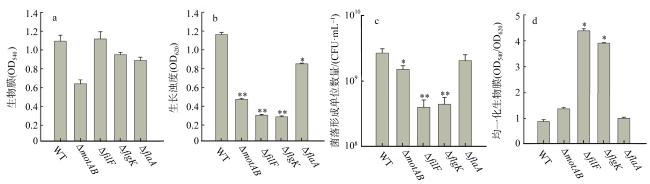
图3 鞭毛缺失对SCSIO 11900生长及生物膜形成的影响野生型SCSIO 11900(wild type, WT)与鞭毛结构基因敲除后的细菌生长7h后, 对96孔板内壁上形成的生物膜进行结晶紫染色, 测定540nm(a)和620nm (b)的吸光度; 同时测定菌落形成单位数量(c), 并对a中的生物膜用b中的生长进行均一化(OD540/OD620) (d)。所有敲除菌株与野生型进行t-test分析, *表示P< 0.05, **表示P< 0.01 Fig. 3 Loss of flagella affects the growth and biofilm formation of SCSIO 11900. Wild type (WT) SCSIO 11900 and flagella destroyed mutant strains grew in LB medium containing 25‰ NaCl for 7h. (a) The biofilm attached on the inner surface of 96 well plate was stained with 0.1% crystal violet and absorbance was determined at 540 nm; (b) determined absorbance at 620 nm and (c) colony forming units (CFU); (d) The biofilm determined in a was normalized by growth in (b). Statistical analysis was performed using student t-test, and all mutant strains were compared to WT. P< 0.05 was shown with one asterisk and P< 0.01was shown with two asterisks |
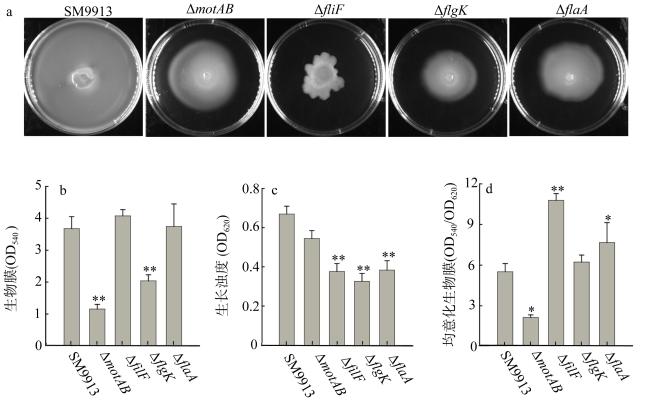
图4 端生鞭毛缺失对SM9913游动性和生物膜形成的影响a. 野生型SM9913与端生鞭毛基因敲除的菌株在游动性平板上游动24h的情况; b. 野生型SM9913与鞭毛结构基因敲除后的细菌生长24h后, 对96孔板内壁上形成的生物膜进行结晶紫染色, 测定540nm的吸光度; c. 测定620nm的吸光度; d. 对b中的生物膜用c中的生长进行了均一化(OD540/OD620)。所有敲除菌株与野生型进行t-test分析, *表示P< 0.05, **表示P< 0.01 Fig. 4 Polar flagella mutant affects motility and biofilm formation of SM9913. (a) Wild type SM9913 and flagella destroyed mutant strains grew in LB medium containing 3‰ agar and 25‰ NaCl for 24h; (b) Wild type SM9913 and flagella destroyed mutant strains grew in LB medium containing 25‰ NaCl in 96-well plate for 24h. The biofilm attached on the inner surface of each well was stained with 0.1% crystal violet and absorbance was determined at 540 nm; (c) Growth was determined as absorbance at 620 nm; (d) The biofilm determined in b was normalized by growth in c (OD540/OD620). Statistical analysis was performed using student t-test, and all mutant strains were compared to WT. P< 0.05 was shown with one asterisk and P< 0.01was shown with two asterisks |
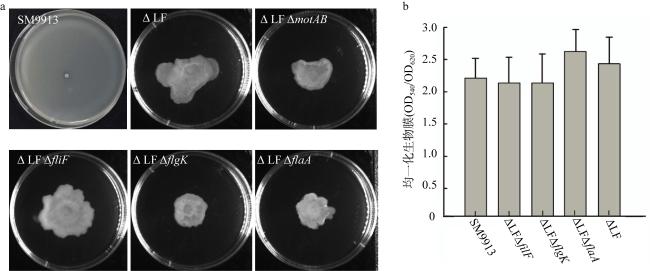
图5 侧生鞭毛缺失对SM9913游动性和生物膜形成的影响a. 野生型SM9913、破坏侧生鞭毛和同时破坏侧生和端生鞭毛的菌株在游动性平板上游动24h后的情况, LF代表具有侧生鞭毛; b. 野生型SM9913与鞭毛破坏的细菌生长24h后, 对生物膜进行了测定 Fig. 5 Loss of lateral flagella inhibits motility of SM9913, but has no effect on biofilm formation. (a)Wild type SM9913 and flagella destroyed mutant strains grew in LB medium containing 25‰ NaCl for 24h; (b) The biofilm attached on the inner surface of 96-well plate was stained with 0.1% crystal violet and normalized by growth |
表3 不同来源的海洋假交替单胞菌均含有端生鞭毛系统, 且部分含有侧生鞭毛系统Tab. 3 Polar flagella exit in P. sp. isolated from different marine environments, and lateral flagella exit in partial strains |
| 菌株 | 基因组NCBI序列号 | 端生鞭毛 | 侧生鞭毛 | 分离环境 | 参考文献 |
|---|---|---|---|---|---|
| P. sp. SCSIO 11900 | JEMJ00000000 | + | - | 中国南海4m水深珊瑚黏液层 | Zeng et al, 2014 |
| P. lipolytica SCSIO 04301 | JDVB00000000 | + | - | 中国南海63m水深沉积物 | Zeng et al, 2014 |
| P. rubra SCSIO 6842 | CP013611/CP013612 | + | - | 孟加拉湾150m深海水 | Li et al, 2016 |
| P. sp. SM9913 | CP001796 | + | + | 冲绳海槽附近1855m沉积物 | Qin et al, 2011 |
| P. tetraodonis UCD-SED8 | LITK01000005 | + | - | 海洋沉积物 | Lee et al, 2015 |
| P. haloplanktis TAC125 | CR954246 | + | - | 大西洋沿岸海水 | Medigue et al, 2005 |
| P. sp. PAMC 22718 | AJTK00000000 | + | + | 喀拉海低温海水 | Park et al, 2012 |
| P. sp. OCN003 | CP011011 | + | - | 夏威夷珊瑚礁区发病珊瑚 | Beurmann et al, 2015 |
| P. distincta ATCC 700518 | NZ_JWIG01000001 | + | - | 白令海的海绵 | Givan et al, 2015 |
| P. elyakovii ATCC 700519 | JWIH00000000 | + | - | 贻贝的体液 | Givan et al, 2015 |
| P. lipolytica UCD-48B | LJTC01000005 | + | - | 海草的叶子 | Alexiev et al, 2016 |
| [1] |
乔毅, 沈辉, 万夕和, 等, 2015. 1株黑鲷致病性假交替单胞菌的鉴定及毒力基因分析[J]. 江苏农业科学, 43(4): 229-233.
|
| [2] |
王枫林, 王秀华, 张宇哲, 等, 2021. 杀鱼假交替单胞菌2515的抗弧菌效果及在对虾养殖中的应用[J]. 中国水产科学, 28(7): 903-913.
|
| [3] |
席宇, 朱大恒, 刘红涛, 等, 2005. 假交替单胞菌及其胞外生物活性物质研究进展[J]. 微生物学通报, 32(3): 108-112.
|
| [4] |
闫咏, 马家海, 许璞, 等, 2002. 1株引起条斑紫菜绿斑病的柠檬假交替单胞菌[J]. 中国水产科学, 9(4): 353-358.
|
| [5] |
张驰, 王劲松, 杨金龙, 等, 2022. 海假交替单胞菌(Pseudoalteromonas marina) pilZ基因缺失抑制厚壳贻贝附着变态[J]. 海洋学报, 44(4): 95-103.
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
BOWMAN J P J M D, 2007. Bioactive compound synthetic capacity and ecological significance of marine bacterial genus Pseudoalteromonas[J]. Marine Drugs, 5(4): 220-241.
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}