
Journal of Tropical Oceanography >
Physiological responses of two soft corals to herbicide acetochlor stress
Copy editor: YAO Yantao
Received date: 2022-08-23
Revised date: 2022-10-06
Online published: 2022-10-11
Supported by
Natural Science Foundation of Hainan Province(321MS0821)
In this paper, using acetochlor (ACT) as the stress environment, the long-term and short-term stress of Sarcophyton sp. and Sinularia sp. with different concentrations of acetochlor were used to study the stress changes in coral physiological indicators. Studies have shown that the chlorophyll a and zooxanthellae contents of succulent soft corals and brachydactyly soft corals increased first and then decreased after different concentrations of acetochlor stress. Superoxide dismutase (SOD) and Catalase (CAT) activity will continue to increase and then decrease and maintain a low level, Alkaline Phosphatase (AKP) activity will first increase and then gradually decrease, Glutathione S-Transferase (GST) activity increased and maintained at a high level. The symbiotic zooxanthellae diversity of succulent soft corals became lower, while the abundance of symbiotic zooxanthellae of Brachydactyly soft corals became lower. The zooxanthellae with high abundance all belonged to group C, while the abundance of group G of Brachydactyly was increased. The abundance of soft coral Cyanobacteria gradually increased and the abundance of Firmicutes gradually decreased after stress. The above studies showed that acetochlor had great changes in coral physiological indicators after the stress of Sarcophyton sp. and Sinularia sp..
Key words: soft coral; acetochlor; zooxanthellae; Physiological indicators
YU Qiang , DONG Xianxian , LI Changqing , ZHANG Xin , ZHAO Guoli , CHIN Yaoxian , WANG Peizheng . Physiological responses of two soft corals to herbicide acetochlor stress[J]. Journal of Tropical Oceanography, 2023 , 42(4) : 133 -145 . DOI: 10.11978/2022183
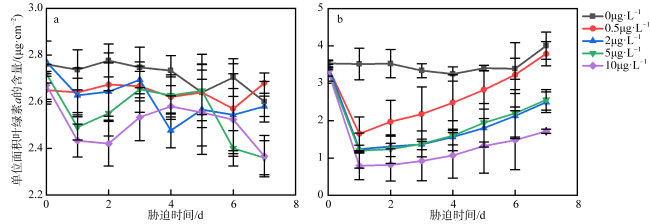
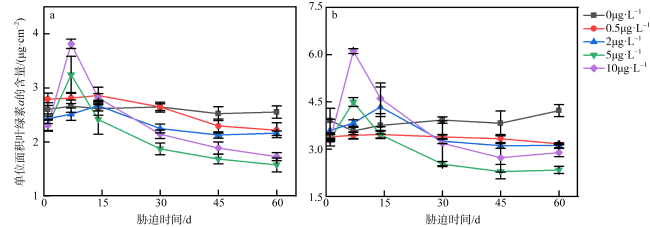
图1 不同浓度乙草胺胁迫下肉质软珊瑚(a)和短指软珊瑚(b)的单位面积叶绿素a浓度变化0μg·L-1表示为空白对照组; 0.5μg·L-1表示软珊瑚在0.5μg·L-1的乙草胺浓度下遭受胁迫; 2μg·L-1表示软珊瑚在2μg·L-1的乙草胺浓度下遭受胁迫; 5μg·L-1表示软珊瑚在5μg·L-1的乙草胺浓度下遭受胁迫; 10μg·L-1表示软珊瑚在10μg·L-1的乙草胺浓度下遭受胁迫 Fig. 1 Changes of chlorophyll a per unit area of the Sarcophyton sp. (a) and Sinularia sp. (b) under different concentrations of acetochlor |
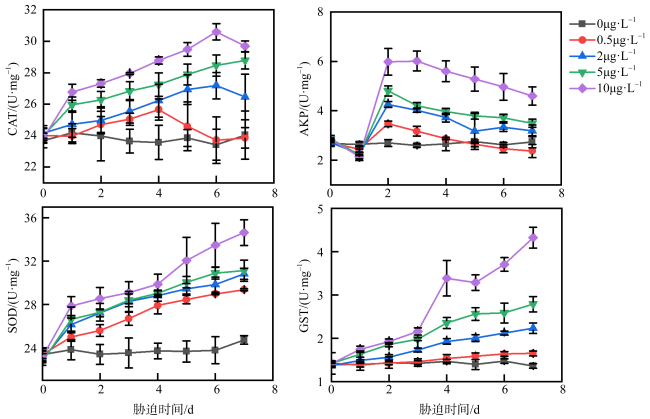
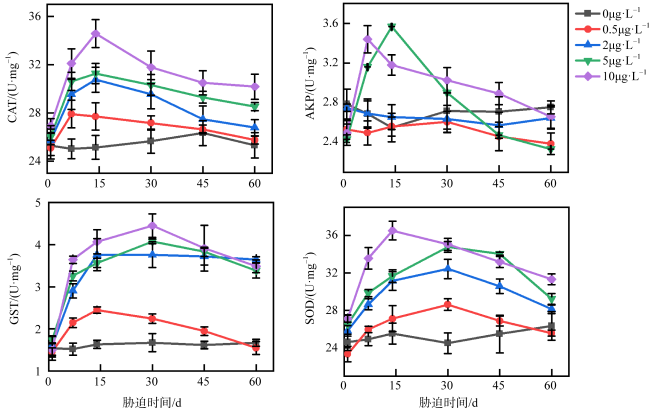
图2 不同浓度乙草胺胁迫下肉质软珊瑚CAT、AKP、SOD、GST活性的变化0μg·L-1表示为空白对照组; 0.5μg·L-1表示肉质软珊瑚在0.5μg·L-1的乙草胺浓度下遭受胁迫; 2μg·L-1表示肉质软珊瑚在2μg·L-1的乙草胺浓度下遭受胁迫; 5μg·L-1表示肉质软珊瑚在5μg·L-1的乙草胺浓度下遭受胁迫; 10μg·L-1表示肉质软珊瑚在10μg·L-1的乙草胺浓度下遭受胁迫 Fig. 2 Changes in the activities of CAT, AKP, SOD, GST enzymes of Sarcophyton sp. under different concentrations of acetoxine stress |
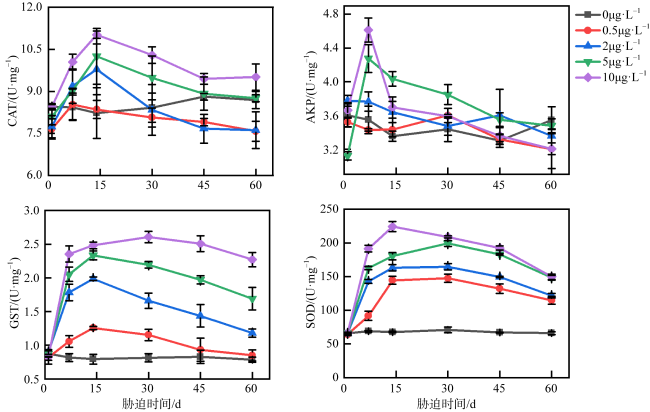
图3 不同浓度乙草胺胁迫下短指软珊瑚CAT、AKP、SOD、GST活性的变化0μg·L-1表示为空白对照组; 0.5μg·L-1表示短指软珊瑚在0.5μg·L-1的乙草胺浓度下遭受胁迫; 2μg·L-1表示短指软珊瑚在2μg·L-1的乙草胺浓度下遭受胁迫; 5μg·L-1表示短指软珊瑚在5μg·L-1的乙草胺浓度下遭受胁迫; 10μg·L-1表示短指软珊瑚在10μg·L-1的乙草胺浓度下遭受胁迫 Fig. 3 Changes in the activities of CAT, AKP, SOD, GST enzymes of Sinularia sp. under different concentrations of acetochlor stress |
表1 短指软珊瑚虫黄藻多样性指数表Tab. 1 Zooxanthellae diversity index of Sinularia sp. |
| 实验类别 | Shannon指数 | Simpson指数 | Ace指数 | Chao指数 | Coverage指数 |
|---|---|---|---|---|---|
| S0S0 | 0.165±0.003 | 0.945±0.001 | 7.4±1.38 | 6.5±0.5 | 1 |
| S24S0.5 | 0.347±0.051* | 0.867±0.021* | 4.5±0.21* | 8.5±0.5 | 1 |
| S24S4 | 0.488±0.156* | 0.798±0.073* | 4.6±0.15 | 7.0±1.0 | 1 |
| S24S8 | 0.279±0.031 | 0.902±0.009 | 4.0±0.14* | 8.5±0.5 | 1 |
| S72S0.5 | 0.273±0.024* | 0.895±0.008* | 3.0±0.95* | 6.5±0.5 | 1 |
| S72S4 | 0.287±0.005* | 0.890±0.001* | 3.0±1.13* | 6.5±0.5 | 1 |
| S72S8 | 0.223±0.025* | 0.922±0.012* | 7.0±1.07 | 7.0±0.6 | 1 |
注: *表示数据差异性显著(P<0.5)。S0S0表示短指软珊瑚空白组; S24S0.5、S24S4、S24S8分别表示在0.5μg·L-1、4μg·L-1、8μg·L-1乙草胺浓度胁迫24h后短指软珊瑚的状态; S72S0.5、S72S4、S72S8分别表示在0.5μg·L-1、4μg·L-1、8μg·L-1乙草胺浓度胁迫72h后短指软珊瑚的状态 |
表2 肉质软珊瑚的虫黄藻多样性指数表Tab. 2 Zooxanthellae diversity index of Sarcophyton sp. |
| 实验类别 | Shannon指数 | Simpson指数 | Ace指数 | Chao指数 | Coverage指数 |
|---|---|---|---|---|---|
| M0M0 | 1.042±0.006 | 0.488±0.009 | 4.0±0.76 | 7.0±1.0 | 1 |
| M24M0.5 | 1.049±0.025 | 0.499±0.002 | 4.5±0.38 | 8.0±0.2 | 1 |
| M24M4 | 1.029±0.008 | 0.503±0.005 | 8.0±1.23 | 8.0±0.1 | 1 |
| M24M8 | 0.759±0.292* | 0.657±0.155* | 9.5±1.5 | 9.0±1.0* | 1 |
| M72M0.5 | 1.066±0.055 | 0.498±0.018 | 4.0±0.28 | 8.0±0.3 | 1 |
| M72M4 | 1.018±0.006 | 0.506±0.005 | 4.0±0.09 | 7.5±0.5 | 1 |
| M72M8 | 0.749±0.283* | 0.660±0.155* | 9.1±0.06* | 8.5±0.5* | 1 |
注: *表示数据差异性显著(P<0.5)。M0M0表示肉质软珊瑚空白组; M24M0.5、M24M4、M24M8分别表示在0.5μg·L-1、4μg·L-1、8μg·L-1乙草胺浓度胁迫24h后肉质软珊瑚的状态; M72M0.5、M72M4、M72M8分别表示在0.5μg·L-1、4μg·L-1、8μg·L-1乙草胺浓度胁迫72h后肉质软珊瑚的状态 |
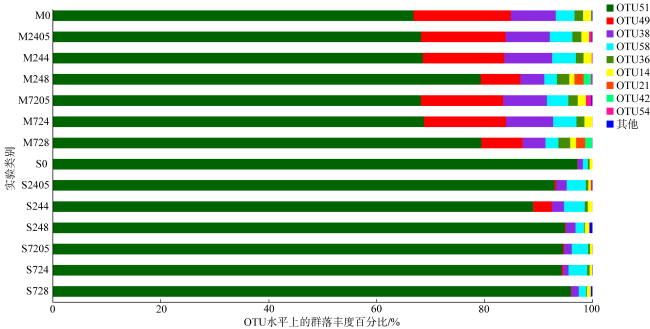
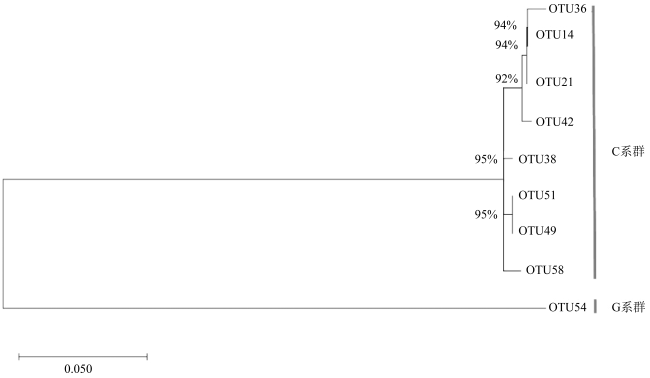
图4 两种珊瑚受到乙草胺胁迫后的共生虫黄藻OTU组成图M0表示肉质软珊瑚空白组; M2405、M244、M248分别表示在0.5μg·L-1、4μg·L-1、8μg·L-1乙草胺浓度胁迫24h后肉质软珊瑚的状态; M7205、M724、M728分别表示在0.5μg·L-1、4μg·L-1、8μg·L-1乙草胺浓度胁迫72h后肉质软珊瑚的状态。S0表示短指软珊瑚空白组; S2405、S244、S248分别表示在0.5μg·L-1、4μg·L-1、8μg·L-1乙草胺浓度胁迫24h后短指软珊瑚的状态; S7205、S724、S728分别表示在0.5μg·L-1、4μg·L-1、8μg·L-1乙草胺浓度胁迫72h后短指软珊瑚的状态。OTU51、OTU49、OTU38、OTU58、OTU36、OTU14、OTU21、OTU42、OTU54代表不同虫黄藻亚种 Fig. 4 Composition of the OTU of two kinds of corals after acetoxaline stress |
表3 肉质软珊瑚细菌多样性指数表Tab. 3 Bacterial diversity index of Sarcophyton sp. |
| 实验类别 | Shannon指数 | Simpson指数 | Ace指数 | Chao指数 | Coverage指数 |
|---|---|---|---|---|---|
| M0M0 | 5.116±0.267 | 0.019±0.003 | 743.1±119.5 | 744.2±121.8 | 1 |
| M24M0.5 | 4.257±0.468* | 0.079±0.023* | 580.0±140.3* | 580.6±137.1* | 1 |
| M24M4 | 3.170±0.046* | 0.221±0.019* | 400.1±26.8* | 401.1±29.5* | 1 |
| M24M8 | 3.595±0.541* | 0.189±0.078* | 763.5±99.5 | 768.6±64.9 | 1 |
| M72M0.5 | 3.932±0.248 | 0.087±0.001* | 519.1±51.0 | 529.9±45.1 | 1 |
| M72M4 | 3.375±0.193* | 0.158±0.004* | 523.1±45.5 | 527.4±42.1 | 1 |
| M72M8 | 5.023±1.039 | 0.032±0.004 | 1065.2±49.7 | 1075.6±65.4 | 1 |
注: *表示数据差异性显著(P<0.5)。M0M0表示肉质软珊瑚空白组; M24M0.5、M24M4、M24M8分别表示在0.5μg·L-1、4μg·L-1、8μg·L-1乙草胺浓度胁迫24h后肉质软珊瑚的状态; M72M0.5、M72M4、M72M8分别表示在0.5μg·L-1、4μg·L-1、8μg·L-1乙草胺浓度胁迫72h后肉质软珊瑚的状态 |
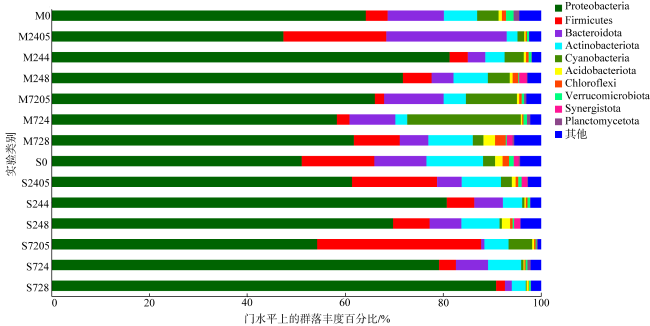
图6 乙草胺胁迫实验中两种珊瑚在门水平上的细菌群落组成图M0表示肉质软珊瑚空白组; M2405、M244、M248分别表示在0.5μg·L-1、4μg·L-1、8μg·L-1乙草胺浓度胁迫24h后肉质软珊瑚的状态; M7205、M724、M728分别表示在0.5μg·L-1、4μg·L-1、8μg·L-1乙草胺浓度胁迫72h后肉质软珊瑚的状态。S0表示短指软珊瑚空白组; S2405、S244、S248分别表示在0.5μg·L-1、4μg·L-1、8μg·L-1乙草胺浓度胁迫24h后短指软珊瑚的状态; S7205、S724、S728分别表示在0.5μg·L-1、4μg·L-1、8μg·L-1乙草胺浓度胁迫72h后短指软珊瑚的状态 Fig. 6 Composition (gate level) of two coral bacterial communities in the acetoxaline stress experiment |
图7 不同浓度乙草胺胁迫下肉质软珊瑚(a)和短指软珊瑚(b)单位面积叶绿素a含量的变化0μg·L-1表示为空白对照组; 0.1μg·L-1表示软珊瑚在0.1μg·L-1的乙草胺浓度下遭受胁迫; 2μg·L-1表示软珊瑚在2μg·L-1的乙草胺浓度下遭受胁迫; 4μg·L-1表示软珊瑚在4μg·L-1的乙草胺浓度下遭受胁迫; 8μg·L-1表示软珊瑚在8μg·L-1的乙草胺浓度下遭受胁迫 Fig. 7 Changes of chlorophyll a per unit area of the Sarcophyton sp. (a) and Sinularia sp. (b) under different concentrations of acetochlor |
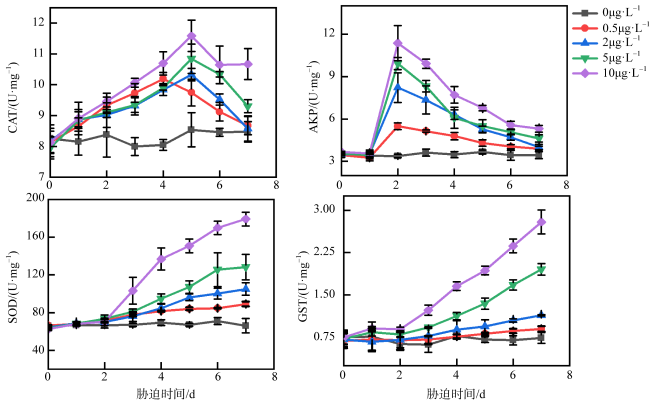
图8 不同浓度乙草胺胁迫下肉质软珊瑚CAT、AKP、SOD、GST活性的变化0μg·L-1表示为空白对照组; 0.1μg·L-1表示肉质软珊瑚在0.1μg·L-1的乙草胺浓度下遭受胁迫; 2μg·L-1表示肉质软珊瑚在2μg·L-1的乙草胺浓度下遭受胁迫; 4μg·L-1表示肉质软珊瑚在4μg·L-1的乙草胺浓度下遭受胁迫; 8μg·L-1表示肉质软珊瑚在8μg·L-1的乙草胺浓度下遭受胁迫 Fig. 8 Changes in the activities of CAT, AKP, SOD, GST enzymes of Sarcophyton sp. under different concentrations of acetoxine stress |
图9 不同浓度乙草胺胁迫下短指软珊瑚CAT、AKP、SOD、GST活性的变化0μg·L-1表示为空白对照组; 0.1μg·L-1表示短指软珊瑚在0.1μg·L-1的乙草胺浓度下遭受胁迫; 2μg·L-1表示短指软珊瑚在2μg·L-1的乙草胺浓度下遭受胁迫; 4μg·L-1表示短指软珊瑚在4μg·L-1的乙草胺浓度下遭受胁迫; 8μg·L-1表示短指软珊瑚在8μg·L-1的乙草胺浓度下遭受胁迫 Fig. 9 Changes in the activities of CAT, AKP, SOD, GST enzymes of Sinularia sp. under different concentrations of acetochlor stress |
| [1] |
晁华, 申玉春, 刘丽, 2016. 氮磷对珊瑚过氧化氢酶和超氧化物歧化酶活性的影响[J]. 中国农学通报, 32(17): 12-19.
|
| [2] |
呼光富, 李忠, 梁宏伟, 等, 2009. 镉对克氏原螯虾肝胰腺触角腺及鳃中SOD和CAT活性的影响[J]. 农业环境科学学报, 28(9): 1806-1811.
|
| [3] |
雷新明, 黄晖, 王华接, 等, 2008. 升温胁迫对珊瑚及其共生藻影响的初步研究[J]. 热带海洋学报, 27(5): 55-59.
|
| [4] |
李俊杰, 李玲, 黄沛玲, 2020. 乙草胺胁迫对水华微囊藻光合生理特性的影响[J]. 生态毒理学报, (5): 1-14.
|
| [5] |
李丽君, 2006. 丁草胺和高效氯氰菊酯对2种海洋微藻的生态毒性效应研究[D]. 广州: 暨南大学.
|
| [6] |
李薪芳, 索亚萍, 楼鸳鸯, 等, 2016. 酰胺类除草剂对铜绿微囊藻的生长影响及氧化损伤效应[J]. 生态毒理学报, 11(1): 239-247.
|
| [7] |
李泽鹏, 2012. 主要环境因子对滨珊瑚的胁迫作用研究[D]. 湛江: 广东海洋大学.
|
| [8] |
林玉锁, 龚瑞忠, 朱忠林, 2000. 农药与生态环境保护[M]. 北京: 化学工业出版社.
|
| [9] |
陆昊, 李卫东, 王沛政, 等, 2019. 长时间微塑料环境暴露对肉芝软珊瑚的影响[J]. 海洋科学, (11): 1-7.
|
| [10] |
陆昊, 刘红岩, 黄秀铭, 等, 2021. 洗涤剂主成分LAS和AEO对软珊瑚氧化应激水平的影响[J]. 海洋环境科学, 40(1): 133-138.
|
| [11] |
乔丹, 2017. 山东沿海贝类中除草剂污染特征及风险评价[D]. 上海: 上海海洋大学.
|
| [12] |
赵美霞, 余克服, 张乔民, 2006. 珊瑚礁区的生物多样性及其生态功能[J]. 生态学报, 26(1): 186-194.
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}