
Journal of Tropical Oceanography >
Effects of ocean acidification on the calcification and gene expression in coral Acropora hyacinthus*
Copy editor: LIN Qiang
Received date: 2022-08-23
Revised date: 2022-12-22
Online published: 2022-12-15
Supported by
National Key Research and Development Program of China(2021YFF0502800)
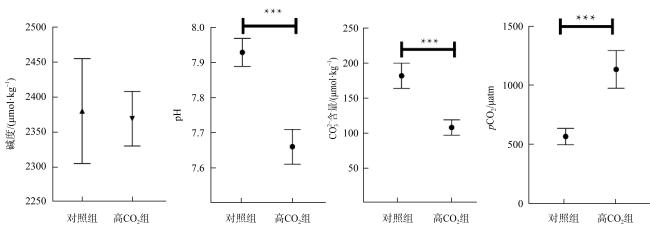
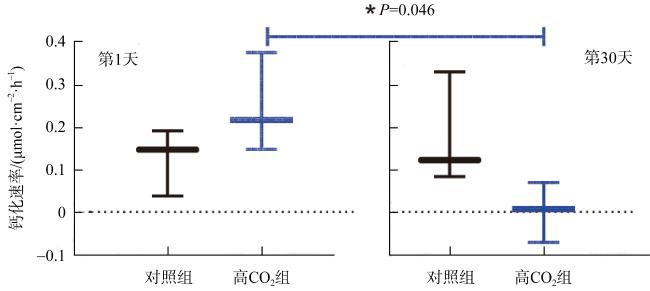
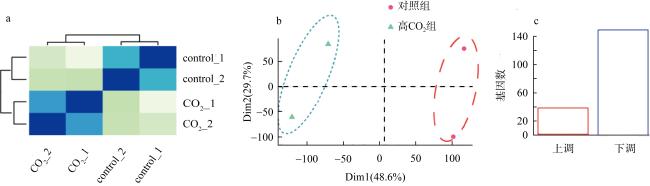
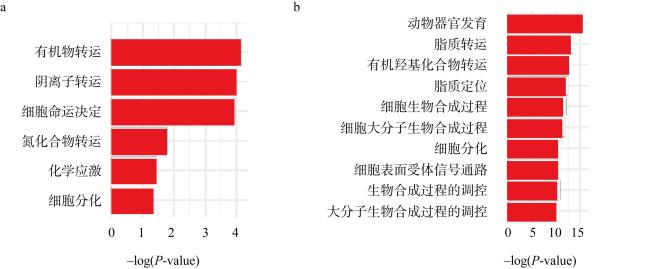
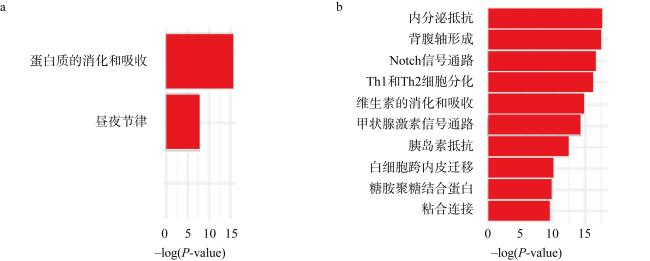
We studied the responses of coral Acropora hyacinthus to the increase of CO2, with a simulated experiment in which corals were cultured in natural seawater as the control group with pCO2 of 565 μatm and in high CO2 treatment with pCO2 of 1135 μatm. The effects of seawater acidification on the calcification and gene expression were investigated. The results showed that under acidification stress, the calcification rate of A. hyacinthus significantly decreased, and the gene expression was greatly affected. As the pathways of organic matter transport, anion transport, chemical stress were up-regulated, and animal organ development, lipid transport, cell biosynthesis, cell surface receptor signal and other pathways were down-regulated. Among the calcification related genes, carbonic anhydrase was down-regulated, and Ca2+-ATPase and calcium ion transport related genes were up-regulated. This experiment showed that high CO2 significantly affected the calcification and gene expression of A. hyacinthus, in which the decrease of calcification was mainly due to the down-regulation of carbonic anhydrase resulting in the decrease of $\mathrm{HCO}_{3}^{-}$.
YUAN Xiangcheng , LIANG Yuxian , SONG Yan , YU Xiaolei , HUANG Hui , ZHOU Weihua . Effects of ocean acidification on the calcification and gene expression in coral Acropora hyacinthus*[J]. Journal of Tropical Oceanography, 2024 , 43(3) : 40 -48 . DOI: 10.11978/2022182
| [1] |
郭亚娟, 周伟华, 袁翔城, 等, 2018. 两种造礁石珊瑚对海水酸化和溶解有机碳加富的响应[J]. 热带海洋学报, 37(1): 57-63.
|
| [2] |
梁宇娴, 俞晓磊, 郭亚娟, 等, 2020. 3种传统方法对不同珊瑚表面积测量的适用性及其校准方法——以3D扫描技术为基准[J]. 热带海洋学报, 39(1): 85-93.
|
| [3] |
朱亚兰, 姚伟, 乐超银, 等, 2006. 甲状旁腺激素促进骨形成的研究进展[J]. 山东医药, 46(7): 79-80. (in Chinese)
|
| [4] |
邹仁林, 2001. 中国动物志, 腔肠动物门, 珊瑚虫纲, 石珊瑚目, 造礁石珊瑚[M]. 北京: 科学出版社 (in Chinese).
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
IPCC, 2022. Special report on the ocean and cryosphere in a changing climate[R]. New York: Cambridge University Press.
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}