
Journal of Tropical Oceanography >
Bioaccumulation characteristics and source apportionment of organophosphate esters in Acanthaster planci from the South China Sea
Received date: 2022-10-19
Revised date: 2022-12-16
Online published: 2022-12-19
Supported by
Guangxi Natural Science Foundation(2020GXNSFDA297005)
Guangxi Laboratory on the Study of Coral Reefs in the South China Sea Independent Project(GXLSCRSCS2021101)
Innovation and Entrepreneurship Training Program of Guangxi University(202210593887)
National Natural Science Foundation of China(42030502)
National Natural Science Foundation of China(42090041)
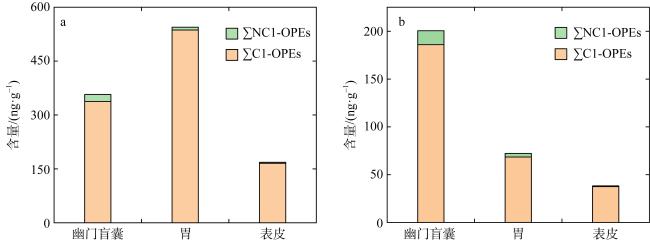
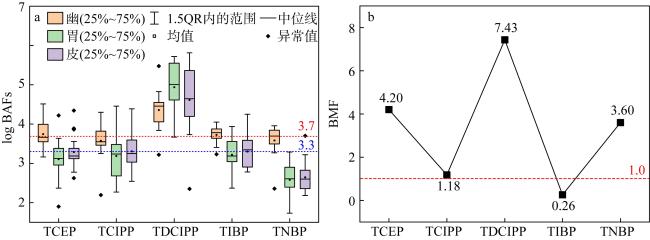
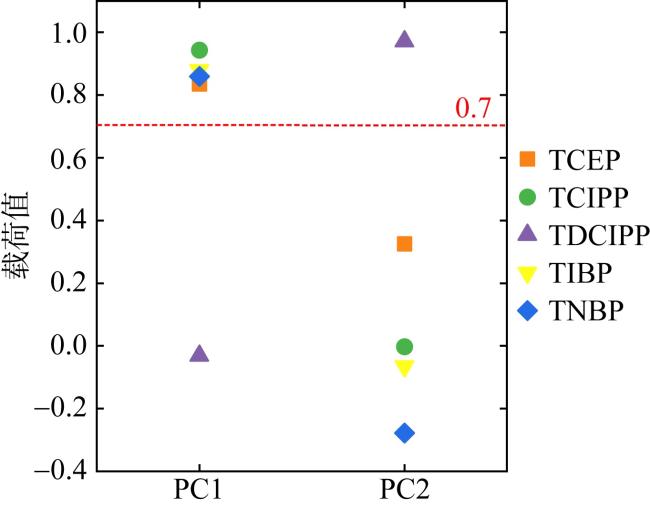
Organophosphate esters (OPEs) is a new type of organic pollutants in the environment, which are ubiquitous in seawater and organisms in the South China Sea. However, the bioaccumulation characteristics of OPEs in the Acanthaster planci of the South China Sea have not been studied. Therefore, in this study, gas chromatography tandem triple quadrupole mass spectrometry (GC-MS/MS) was used to analyze the bioaccumulation characteristics and sources of 11 typical OPEs in the Acanthaster planci in coral reef regions of the South China Sea. The results showed that chlorinated OPEs were ubiquitous in the tissues of Acanthaster planci, accounting for 95% ~ 98% of the total concentrations of 11 OPEs (∑11OPEs). ∑11OPEs (dry weight) in Acanthaster planci showed significant differences among tissues, and ∑11OPEs in the pyloric caecum (60.2 ~ 1130 ng·g-1) and stomach (68.2 ~ 1181 ng·g-1) were significantly higher than those in the epidermis (9.31 ~ 569 ng·g-1) (p < 0.05), pyloric caecum and stomach played an important role in the accumulation of OPEs in the Acanthaster planci. Tri (1,3-dichloro-2-isopropyl) phosphate (TDCIPP) showed strong bioaccumulation ability in the tissues of Acanthaster planci (The bioconcentration factor BAFs of TDCIPP in the Acanthaster planci exceeds 5000 L·kg-1). Tris (2-chloroethyl) phosphate (TCEP), tris (2-chloroisopropyl) phosphate (TCIPP), TDCIPP, and Tri-n-butyl phosphate (TNBP) showed a biomagnification effect in the Acanthaster planci, indicating that these four OPEs can be accumulated by feeding on coral. OPEs in the Acanthaster planci of the South China Sea may mainly come from the release of fishery activities, island and reef buildings, and domestic wastes from surrounding countries, and a small part may come from the release of ship transportation.
WANG Chenyan , SHI Jingwen , YAN Annan , KANG Yaru , WANG Yuxuan , QIN Suli , HAN Minwei , ZHANG Ruijie , YU Kefu . Bioaccumulation characteristics and source apportionment of organophosphate esters in Acanthaster planci from the South China Sea[J]. Journal of Tropical Oceanography, 2023 , 42(5) : 30 -37 . DOI: 10.11978/2022223
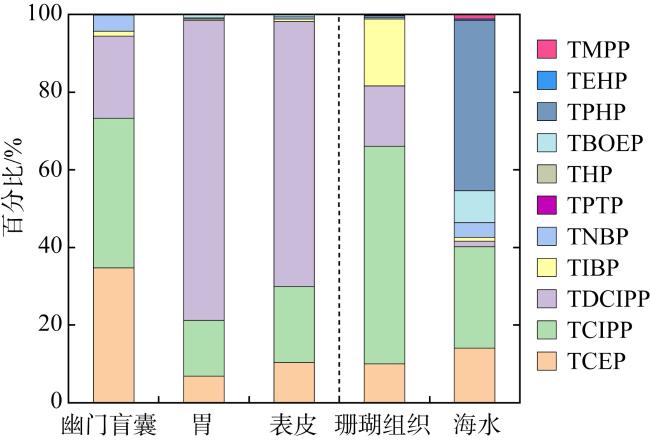
表1 南海长棘海星不同组织(干重)中OPEs的检出率和含量水平(单位: ng·g-1)Tab. 1 Detection rates and concentrations of OPEs in different tissues of Acanthaster planci (dry weight) from the South China Sea (unit: ng·g-1) |
| OPEs | 幽门盲囊(n=16) | 胃(n=17) | 表皮(n=24) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 检出率/% | 范围 | 平均值± 标准差 | 中位数 | 检出率/% | 范围 | 平均值± 标准差 | 中位数 | 检出率/% | 范围 | 平均值± 标准差 | 中位数 | |
| TCEP | 100 | 14.6~532 | 124±150 | 67.3 | 100 | 4.68~153 | 37.2±41.2 | 20.2 | 100 | 3.32~61.4 | 17.4±15.3 | 11.4 |
| TCIPP | 100 | 2.87~552 | 138±148 | 87.5 | 100 | 6.22~485 | 78.2±107 | 45.5 | 100 | 5.07~115 | 33.1±28.9 | 22.7 |
| TDCIPP | 81 | nd~319 | 75.3±105 | 34.6 | 88 | nd~1086 | 421±353 | 397 | 100 | 0.19~486 | 115±147 | 39.2 |
| TIBP | 100 | 0.50~11.3 | 4.61±2.65 | 4.70 | 82 | nd ~5.54 | 1.74±1.63 | 1.13 | 88 | nd~2.60 | 1.11±0.76 | 1.13 |
| TNBP | 88 | nd~35.7 | 15.0±10.2 | 12.8 | 76 | nd ~4.82 | 1.45±1.39 | 1.28 | 79 | nd~2.95 | 0.92±0.78 | 0.85 |
| TPTP | 0 | nd | nd | nd | 0 | nd | nd | nd | 8 | nd~0.57 | 0.04±0.13 | nd |
| THP | 0 | nd | nd | nd | 0 | nd | nd | nd | 0 | nd | nd | nd |
| TBOEP | 13 | nd ~5.33 | 0.46±1.35 | nd | 18 | nd~50.9 | 4.69±12.6 | nd | 17 | nd~6.73 | 0.88±2.00 | nd |
| TPHP | 0 | nd | nd | nd | 0 | nd | nd | nd | 0 | nd | nd | nd |
| TEHP | 13 | nd~0.40 | 0.03±0.10 | nd | 12 | nd~2.08 | 0.24±0.67 | nd | 8 | nd~1.47 | 0.12±0.39 | nd |
| TMPP | 0 | nd | nd | nd | 0 | nd | nd | nd | 0 | nd | nd | nd |
| ∑11OPEs | 60.2~1130 | 357±298 | 262 | 68.2~1181 | 544±409 | 487 | 9.31~569 | 169±161 | 97.0 | |||
注: nd表示未检出 |
表2 南海珊瑚组织(干重)(单位: ng·g-1)和海水(单位: ng·L-1)中OPEs的检出率和含量水平Tab. 2 Detection rates and concentrations of OPEs in coral tissues (dry weight) (unit: ng·g-1) and seawater (unit: ng·L-1) from the South China Sea |
| OPEs | 珊瑚组织(n=9) | 海水(n=2) | ||||||
|---|---|---|---|---|---|---|---|---|
| 检出率/% | 范围 | 平均值±标准差 | 中位数 | 检出率/% | 范围 | 平均值±标准差 | 中位数 | |
| TCEP | 22 | nd ~96.5 | 11.9±30.1 | nd | 100 | 1.78~2.60 | 2.19±0.41 | 2.19 |
| TCIPP | 33 | nd ~278 | 66.9±105 | nd | 100 | 2.45~5.61 | 4.03±1.58 | 4.03 |
| TDCIPP | 11 | nd~166 | 18.5±52.2 | nd | 100 | 0.20~0.26 | 0.23±0.03 | 0.23 |
| TIBP | 78 | nd ~87.0 | 20.6±25.3 | 19.0 | 100 | 0.08~0.21 | 0.15±0.06 | 0.15 |
| TNBP | 11 | nd~4.86 | 0.54±1.53 | nd | 100 | 0.50~0.68 | 0.59±0.09 | 0.59 |
| TPTP | 0 | nd | nd | nd | 0 | nd | nd | nd |
| THP | 0 | nd | nd | nd | 0 | nd | nd | nd |
| TBOEP | 0 | nd | nd | nd | 100 | 0.18~2.38 | 1.28±1.10 | 1.28 |
| TPHP | 11 | nd~4.73 | 0.53±1.49 | nd | 100 | 5.16~8.42 | 6.79±1.63 | 6.79 |
| TEHP | 11 | nd~2.97 | 0.33±0.93 | nd | 100 | 0.05~0.08 | 0.06±0.02 | 0.06 |
| TMPP | 0 | nd | nd | nd | 100 | 0.09~0.27 | 0.18±0.09 | 0.18 |
| ∑11OPEs | nd~461 | 119±163 | 19.8 | 15.0~16.0 | 15.5±0.45 | 15.5 | ||
注: nd表示未检出 |
图2 南海长棘海星不同组织中∑11OPEs的含量差异a. 南海长棘海星不同组织中∑11OPEs (干重)的含量差异; b. 南海长棘海星不同组织中∑11OPEs (脂重)的含量差异 Fig. 2 ∑11OPEs in different tissues of Acanthaster planci from the South China Sea. (a) Differences in the content of ∑11OPEs (dry weights) in different tissues of Acanthaster planci from the South China Sea; (b) Differences in the content of ∑11OPEs (lipid weights) in different tissues of Acanthaster planci from the South China Sea |
图3 南海长棘海星不同组织中OPEs的生物富集因子常用对数(log BAFs, a)和生物放大因子(BMF, b)图a中的25%~75%的含义是指代从小到大排序处于25%至75%区间的数据, 以箱体长度表示, 一定程度上表示数据的波动程度 Fig. 3 The Commonly used logarithms of the bioaccumulation factors (log BAFs, a) and the biomagnification factors (BMF, b) of OPEs in different tissues of Acanthaster planci from the South China Sea |
| [1] |
李厚金, 李淑青, 王丰, 等, 2009. 长棘海星的体内脂肪酸成分及其生物学意义分析[J]. 中山大学学报(自然科学版), 48(5): 55-60.
|
| [2] |
李元超, 梁计林, 吴钟解, 等, 2019. 长棘海星的暴发及其防治[J]. 海洋开发与管理, 36(8): 9-12.
|
| [3] |
廖梓聪, 李会茹, 杨愿愿, 等, 2022. 有机磷酸酯(OPEs)的环境污染特征、毒性和分析方法研究进展[J]. 环境化学, 41(4): 1193-1215.
|
| [4] |
吕佳佩, 张振飞, 刘杨, 等, 2020. 太湖重点区域多介质水体中有机磷酸酯的分布特征及来源解析[J]. 环境科学, 41(12): 5438-5447.
|
| [5] |
陆运涛, 赵赛, 仇雁翎, 等, 2018. 塘西河沉积物中有机磷酸酯的污染现状及来源解析[J]. 环境污染与防治, 40(5): 604-608.
|
| [6] |
彭晓彤, 钟广法, 2020. 南海深海塑料垃圾污染[J]. 科技导报, 38(18): 99-104.
|
| [7] |
张文萍, 张振飞, 郭昌胜, 等, 2021. 环太湖河流及湖体中有机磷酸酯的污染特征和风险评估[J]. 环境科学, 42(4): 1801-1810.
|
| [8] |
赵思佳, 张媛媛, 余克服, 等, 2022. 南海珊瑚礁区棘冠海星重金属含量及其生物积累特征分析[J]. 海洋环境科学, 41(4): 579-585.
|
| [9] |
朱金财, 马玉欣, 蔡明红, 2021. 海洋环境PAHs研究进展: 来源、分布及生物地球化学过程[J]. 海洋环境科学, 40(3): 468-476.
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
EUROPEAN CHEMICALS AGENCY, 2017. Guidance on information requirements and chemical safety assessment: Chapter R. 11: PBT/vPvB assessment (Version 3.0)[L].
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}