
Journal of Tropical Oceanography >
Swarms of pelagic gelatinous tunicates and their roles in marine biological carbon pump*
Received date: 2022-11-08
Revised date: 2022-11-27
Online published: 2023-02-13
Supported by
National Natural Science Foundation of China(31971432)
National Natural Science Foundation of China(41976112)
National Natural Science Foundation of China(32171548)
Mucus-feeding pelagic tunicates are widely distributed in the open ocean and coastal waters, which have very high rates of reproduction, and the ability to form massive blooms. In this review, we provide a qualitative overview of the combination of high particle consumption and swarms of pelagic tunicates led to accumulate at the ocean floor as jelly-POM (particulate organic matter), substantial contributions to vertical export fluxes. As well as the swarms in relation to environmental drivers and unique life-history adaptation, its role in pelagic-benthic coupling, structure and energy flow of marine food web by selecting feeding are reviewed. Because pelagic tunicates have high filtering rates and efficiencies and can reach great abundances, they have the potential to remove a significant fraction of shelf water column primary production, are exported via mucous aggregates, fecal pellets, and jelly falls sinking to depth or restructure shelf pelagic food webs. The study of jelly-falls represents a major challenge in the understanding of the biological pump mainly due to technical/sampling hurdles, the ecological significance of pelagic tunicate blooms, for instance, the rate of this downward carbon flux may be further enhanced through in-situ observations on pelagic tunicates’ diel vertical migrations together with quantitative net catches. Future work should pay more attention to the coupling between fine scales of hydrodynamic grazing and breeding rates under in situ conditions, and link bloom impact on carbon cycling to more dimensions of the physical, chemical, and biological ocean environment, in order to more accurately assess the efficiency of the jelly carbon pump and its response to global change.
Key words: pelagic tunicates; swarms or aggregations; jelly-carbon pump; food web
TAN Yehui , LAI Yanjiao , LIAN Xiping , LIU Jiaxing , KE Zhixin , LI Kaizhi . Swarms of pelagic gelatinous tunicates and their roles in marine biological carbon pump*[J]. Journal of Tropical Oceanography, 2023 , 42(5) : 178 -193 . DOI: 10.11978/2022239
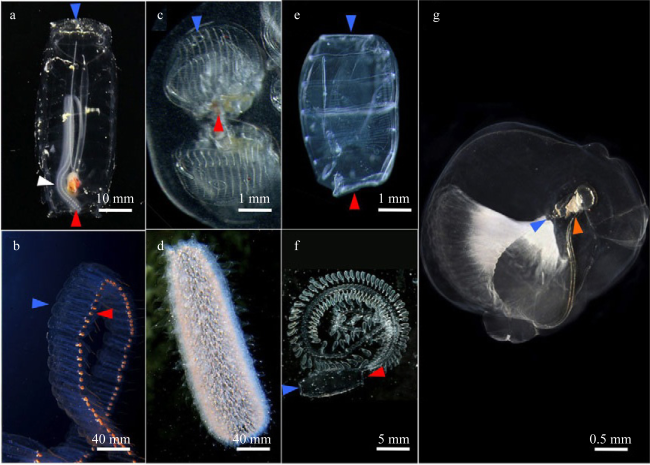
图1 浮游被囊动物成体形态[据Lemaire (2011)修改]a, b: 纽鳃樽; c, d: 火体虫; e, f: 海樽; g. 有尾类。白色箭头表示无性繁殖个体上存在初期的有性繁殖个体群; 红色箭头表示口腔; 蓝色箭头表示心房虹吸管 Fig. 1 The adult morphological features of planktonic tunicates. (a, b) Salp; (c, d) Pyrosome; (e, f) Doliolid; (g) Appendicularian; white arrowheads indicate asexual zooid with a nascent chain of sexual zooids; blue and red arrowheads indicate oral and atrial siphons respectively, modified from Lemaire (2011) |
*感谢丁翔同学在文献编辑方面的帮助。
| [1] |
李超伦, 孙松, 张光涛, 等, 2000. 南极普里兹湾临近海域夏季纽鳃樽对浮游植物的摄食研究[J]. 极地研究, 12(2): 97-104.
|
| [2] |
黄邦钦, 邱勇, 陈纪新, 2019. 海洋生物泵研究的若干新进展与展望[J]. 应用海洋学学报, 38(4): 474-483.
|
| [3] |
谭激扬, 黄良民, 谭烨辉, 等, 2013. 水团对吕宋海峡浮游植物群落结构的影响[J]. 海洋学报, 35(6): 178-189.
|
| [4] |
张金标, 黄将修, 连光山, 2003a. 台湾南湾区秋末冬初海樽类的种类组成和数量分布[J]. 海洋通报, 22(6): 9-16.
|
| [5] |
张金标, 连光山, 王云龙, 等, 2003b. 台湾海峡东部海域海樽类被囊动物的分布[J]. 台湾海峡, 22(3): 279-285.
|
| [6] |
张武昌, 张芳, 王克, 2001. 海洋浮游动物粪便通量[J]. 地球科学进展, 16(1): 113-119.
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|
| [122] |
|
| [123] |
|
| [124] |
|
| [125] |
|
| [126] |
|
| [127] |
|
| [128] |
|
| [129] |
|
| [130] |
|
| [131] |
|
| [132] |
|
| [133] |
|
| [134] |
|
| [135] |
|
| [136] |
|
| [137] |
|
| [138] |
|
| [139] |
|
| [140] |
|
| [141] |
|
| [142] |
|
| [143] |
|
| [144] |
|
| [145] |
|
| [146] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}