
Journal of Tropical Oceanography >
Transcriptomic analysis of fatty acid metabolism in the Thalassiosira pseudonana under low salinity stress
Received date: 2022-12-01
Revised date: 2023-01-12
Online published: 2023-02-22
Supported by
Qingdao National Laboratory of Marine Science and Technology(2021QNLM040001)
National Natural Science Foundation of China(41976133)
The expression of fatty acid metabolism-related genes of Thalassiosira pseudonana under different salinities and different growth stages was analyzed by transcriptomic sequencing in this study. A total of 40 coding genes that are related to the fatty acid metabolisms were retrieved from this study with distinct expression levels at different salinity concentrations. The results showed that the expression levels of genes related to fatty acid biosynthesis and elongation (such as ACC1, armB) in T. pseudonana cells were significantly higher at the second and fourth days compared to those at the first day across the salinity gradients, while the expression levels of genes related to fatty acid degradation (such as ACADM, ECI1) did not change significantly. The expression of genes related to fatty acid biosynthesis and elongation in the experimental group were differentially expressed compared to the control group (for example, the expression levels of KASⅠ, ACAA2 and other genes, which play an important role, increased significantly, but a few of them decreased or changed slightly), although the fatty acid degradation related genes were significantly increased in the experimental group. This study improves our understanding of the survival and adaptation strategies of diatoms in the offshore euryhaline environments, and supports the exploration of outbreak and vanishment of harmful algal blooms.
Key words: Thalassiosira pseudonana; fatty acid metabolism; RNA-seq; salinity stress
SUN Wenjie , LI Jiamin , WANG Hualong , MI Tiezhu , ZHEN Yu . Transcriptomic analysis of fatty acid metabolism in the Thalassiosira pseudonana under low salinity stress[J]. Journal of Tropical Oceanography, 2023 , 42(5) : 92 -103 . DOI: 10.11978/2022250
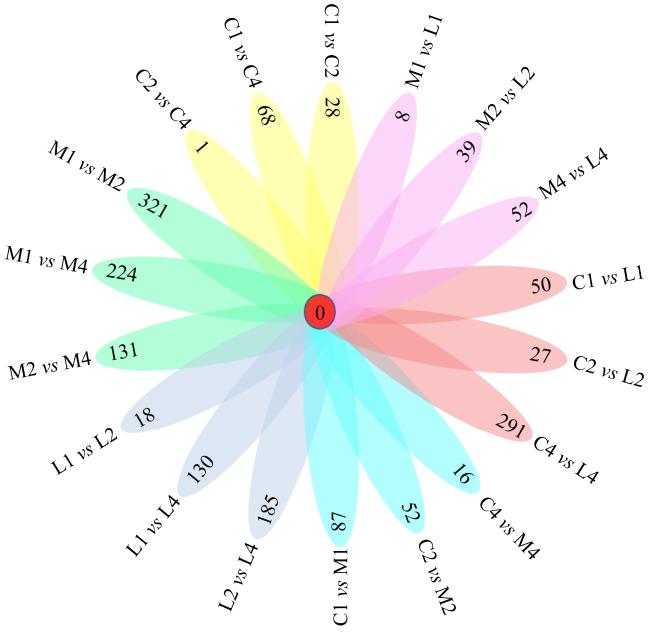
图1 不同比较组间的共有差异表达基因C1、C2和C4分别为对照组第1天、第2天和第4天3个试验组; M1、M2和M4分别为中盐组第1天、第2天和第4天3个试验组; L1、L2和L4分别为高盐组第1天、第2天和第4天3个试验组。每一个椭圆形中的数字表示两组间共有的差异表达基因的数目, 中间红色的“0”代 Fig. 1 Shared expressed genes between two groups. C1/C2/C4: the control group on day one, day two and day four; M1/M2/M4: the middle salinity group on day one, day two and day four; L1/L2/L4: the low salinity group on day one, day two and day four. Number in each oval represents the count of shared expressed genes between two groups |
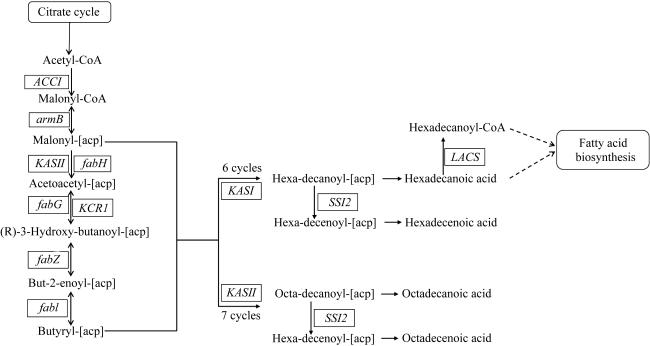
图3 脂肪酸的生物合成及延伸途径Citrate cycle: 柠檬酸循环; Acetyl-CoA: 乙酰辅酶A; Malonyl-CoA: 丙二酰-CoA; Malonyl-[acp]: 丙二酸单酰-ACP; Acetoacetyl-[acp]: 乙酰乙酰-ACP; (R)-3-Hydroxy-butanoyl-[acp]: 3-羟基丁酰基-ACP; But-2-enoyl-[acp]: 丁-2-烯酰基-ACP; Butyryl-[acp]: 丁酰-ACP; Hexa-decanoyl-[acp]: 十六酰-ACP; Hexa-decenoyl-[acp]: 十六烯酰基-ACP; Hexadecenoic acid: 十六烯酸; Hexadecanoyl-CoA: 十六酰-CoA; Octa-decanoyl-[acp]: 十八酰-ACP; Octa-decanoyl-[acp]: 十八烷酰基-ACP; Octadecenoic acid: 十八烯酸; Fatty acid biosynthesis: 脂肪酸生物合成; ACC1: 乙酰-CoA羧化酶基因; armB: 丙二酸单酰CoA:ACP转酰酶基因; fabH: β-酮酰-ACP合酶Ⅲ基因; fabG: β-酮酰基-ACP还原酶基因; KCR1: β-酮脂酰-辅酶A还原酶基因; fabZ: β-羟酰-ACP脱水酶基因; fabI: 烯酰-ACP还原酶基因; KASⅠ: β-酮酰基-ACP合酶Ⅰ基因; KASII: β-酮酰基-ACP合酶Ⅱ基因; LACS: 长链酰基辅酶A合成酶基因; SSI2: 酰基-ACP去饱和酶基因 Fig. 3 Biosynthesis and elongation of fatty acids |
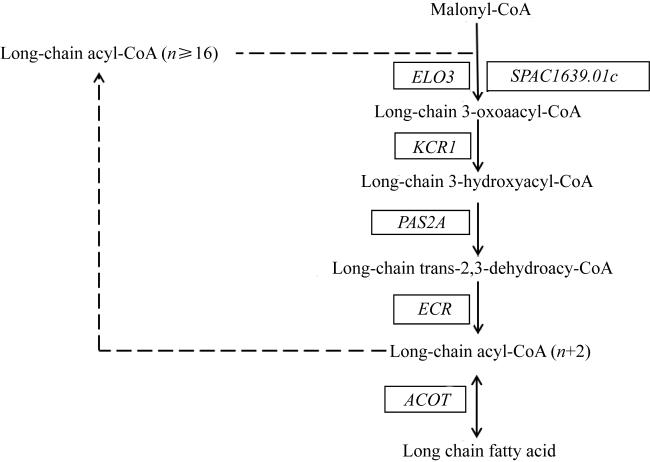
图4 长链脂肪酸的延伸途径Malonyl-CoA: 丙二酸单酰-CoA; Long-chain acyl-CoA: 长链脂酰基-CoA; Long-chain 3-oxoaacyl-CoA: 长链3-乙二酰-CoA; Long-chain 3-hydroxyacyl-CoA: 长链3-脱氢酰基-CoA; Long-chain trans-2,3-dehydroacyl-CoA: 长链2,3-反式脱氢酰基-CoA; Long-chain fatty acid: 长链脂肪酸; ELO3: 脂肪酸延长酶3基因; SPAC1639.01c: 超长链脂肪酸延伸蛋白基因; KCR1: β-酮酰-CoA还原酶1基因; PAS2A: 超长链羟酰-CoA脱水酶基因; ECR: 烯酰-CoA还原酶基因; ACOT: 酰基-CoA硫酯酶基因 Fig. 4 Long chain fatty acids elongation pathway |
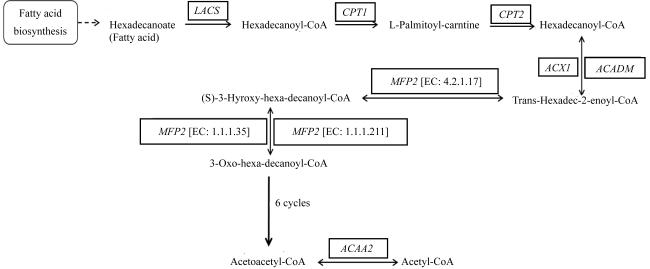
图5 脂肪酸的降解途径Fatty acid biosynthesis: 脂肪酸生物合成; Hexadecanoate: 十六烯酸; Hexadecanoyl-CoA: 十六酰-CoA; L-Palmitoyl-carntine: L-棕榈酰肉碱; Trans-Hexadec-2-enoyl-CoA: 反式-十六烷基-2-烯酰-CoA; (S)-3-Hydroxy-hexa-decanoyl-CoA: (S)-3-羟基十六烷酰-CoA; 3-Oxo-hexa-decanoyl-CoA: 3-氧代十六烷酰-CoA; Acetoacetyl-CoA: 乙酰乙酰-CoA; Acetyl-CoA: 乙酰辅酶A; LACS: 长链酰基辅酶A合成酶基因; CPT: 肉碱棕榈酰转移酶基因; ACX1: 酰基-CoA氧化酶基因; ACADM: 特异性酰基-CoA脱氢酶基因; MFP2: 3-羟酰-CoA脱氢酶基因; ACAA2: 乙酰-CoA酰基转移酶2基因 Fig. 5 Fatty acids degradation pathway |
表1 假微型海链藻转录组中与脂肪酸代谢相关的酶以及对应的转录本Tab. 1 Enzymes and transporters which is associated with fatty acid metabolism in the T. pseudonana transcriptome |
| 信号通路 | 酶(蛋白)名称 | EC编号 | 基因ID | 基因名称 |
|---|---|---|---|---|
| 脂肪酸的 生物合成 | 乙酰-CoA羧化酶(Acetyl-CoA carboxylase) | [EC:6.4.1.2] | THAPS_6770 | ACC1 |
| THAPSDRAFT_12234 | ACC1 | |||
| 丙二酸单酰CoA转移酶([acyl-carrier-protein] S-malonyltransferase) | [EC:2.3.1.39] | THAPSDRAFT_5219 | armB | |
| β-酮酰-ACP合酶Ⅱ (3-oxoacyl-[acyl-carrier-protein] synthase Ⅱ) | [EC:2.3.1.179] | THAPSDRAFT_961 | KASⅡ | |
| THAPSDRAFT_12152 | KAS | |||
| THAPSDRAFT_5021 | KASⅠ | |||
| THAPSDRAFT_262879 | KAS | |||
| β-酮酰-ACP合酶Ⅲ (3-oxoacyl-[acyl-carrier-protein] synthase Ⅲ) | [EC:1.1.1.180] | THAPSDRAFT_270141 | fabH | |
| 长链酰基-CoA合成酶(Long chain acyl-CoA synthetase) | [EC:6.2.1.3] | THAPS_263105 | AAE16 | |
| THAPSDRAFT_11953 | LACS7 | |||
| THAPSDRAFT_270334 | AAE15 | |||
| THAPSDRAFT_29867 | LACS7 | |||
| THAPSDRAFT_13156 | LACS8 | |||
| THAPSDRAFT_25042 | LACS7 | |||
| THAPSDRAFT_262242 | LACS6 | |||
| β-羟酰-ACP脱水酶(3-hydroxyacyl-[acyl-carrier-protein] dehydratase) | [EC:4.2.1.59] | THAPSDRAFT_34681 | fabZ | |
| 烯酰-ACP还原酶(Enoyl-[acyl-carrier protein] reductase I) | [EC:1.3.1.9] | THAPSDRAFT_32860 | fabI | |
| β-酮酰-ACP还原酶(3-oxoacyl-[acyl-carrier-protein] reductase) | [EC:1.1.1.100] | THAPSDRAFT_270321 | fabG | |
| 酰基-ACP去饱和酶(Stearoyl-[acyl-carrier-protein]9-desaturase 2) | [EC:1.14.19.2] | THAPSDRAFT_bd1192 | SSI2 | |
| 脂肪酸的 延伸 | 超长链脂肪酸延伸蛋白(Elongation of fatty acids protein) | [EC:2.3.1.199] | THAPSDRAFT_3741 | ELO3 |
| THAPSDRAFT_728 | SPAC1639.01c | |||
| THAPSDRAFT_22362 | KCS9 | |||
| 乙酰-CoA转移酶(Acetyl-CoA acetyltransferase) | [EC:2.3.1.16] | THAPSDRAFT_34809 | ACAA2 | |
| 超长链酮酰-CoA还原酶(Very-long-chain 3-oxoacyl-CoA reductase) | [EC:1.1.1.330] | THAPSDRAFT_22650 | KCR1 | |
| THAPS_35459 | ACLA_070510 | |||
| 超长链羟酰-CoA脱水酶(Very-long-chain (3R)-3-hydroxyacyl-CoA dehydratase) | [EC:4.2.1.134] | THAPS_6781 | PAS2A | |
| 烯酰-CoA水合酶(Probable enoyl-CoA hydratase 1) | [EC:4.2.1.17] | THAPSDRAFT_34511 | ECHS1 |
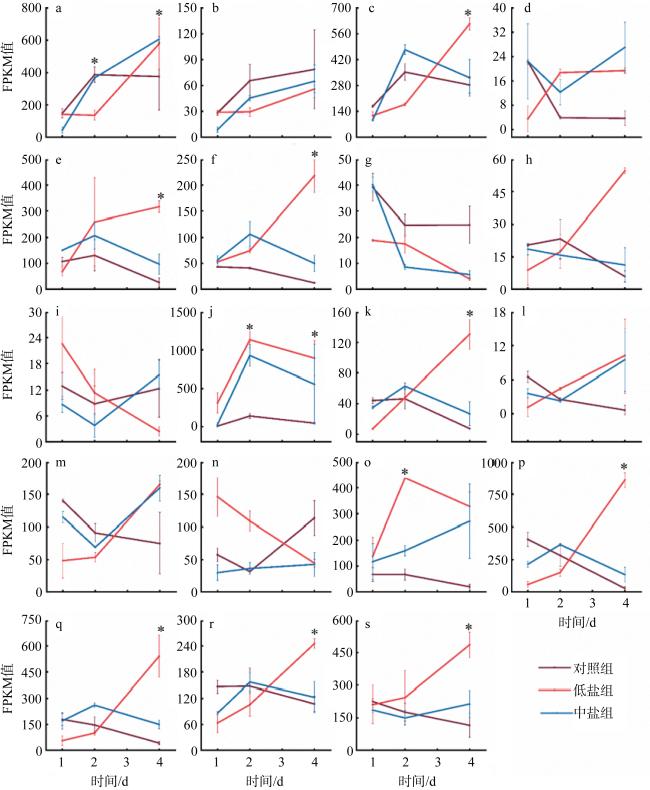
图6 不同盐度条件下假微型海链藻与脂肪酸合成有关的基因差异表达a. THAPS_6770; b. THAPSDRAFT_12234; c. THAPSDRAFT_5219; d. THAPSDRAFT_961; e. THAPSDRAFT_12152; f. THAPSDRAFT_5021; g. THAPSDRAFT_262879; h. THAPSDRAFT_270141; i. THAPSDRAFT_263105; j. THAPSDRAFT_11953; k. THAPSDRAFT_270334; l. THAPSDRAFT_29867; m. THAPSDRAFT_13156; n. THAPSDRAFT_25042; o. THAPSDRAFT_262242; p. THAPSDRAFT_34681; q. THAPSDRAFT_32860; r. THAPSDRAFT_270321; s. THAPSDRAFT_bd1192。上下线代表FPKM值的误差棒; *代表同一时间不同组别或者是同一组别在不同时间的FPKM值具有显著差异 Fig. 6 Gene expression involving in fatty acid synthesis in the T. pseudonana under different salinity conditions |
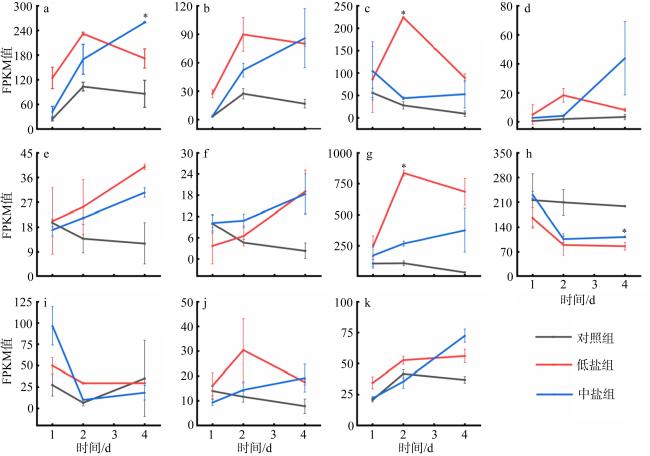
图7 不同盐度条件下假微型海链藻与脂肪酸延伸有关的基因差异表达a. THAPSDRAFT_3741; b. THAPSDRAFT_728; c. THAPSDRAFT_23362; d. THAPSDRAFT_34809; e. THAPSDRAFT_22650; Fig. 7 Gene expression involving in the fatty acid elongation in the T. pseudonana under different salinity conditions |
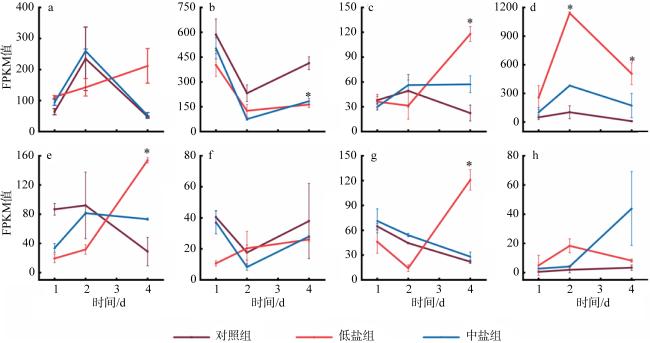
图8 不同盐度条件下假微型海链藻与脂肪酸降解有关的基因差异表达a. THAPSDRAFT_23830; b. THAPSDRAFT_28651; c. THAPSDRAFT_26365; d. THAPSDRAFT_34511; e. THAPSDRAFT_263878; Fig. 8 Gene expression involving in the fatty acid degradation in the T. pseudonana under different salinity conditions |
| [1] |
曹月蕾, 2021. 集胞藻PCC6803类胡萝卜素和脂肪酸代谢途径关键基因功能研究[D]. 济南: 山东师范大学.
|
| [2] |
高彦龙, 吴玉霞, 张仲兴, 等, 2022. 苹果ELO家族基因鉴定及其在低温胁迫下的表达分析[J]. 园艺学报, 49(8): 1621-1636.
|
| [3] |
高羽荞, 闫博巍, 赵莹, 等, 2018. 碱蓬转录组分析及油脂合成相关基因表达模式[J]. 中国油料作物学报, 40(6): 801-811.
|
| [4] |
黄璐, 2020. 海洋硅藻假微型海链藻休眠细胞形成过程的应激响应与长期适应机制[D]. 厦门: 厦门大学.
|
| [5] |
刘淑雅, 陈楠生, 2021. 胶州湾海域浮游植物和赤潮物种的生物多样性研究进展[J]. 海洋科学, 45(4): 170-188.
|
| [6] |
刘璇, 张莹, 李爱芬, 2022. 活性氧介导微藻脂质积累的研究现状及进展[J]. 植物生理学报, 58(7): 1201-1211.
|
| [7] |
聂煜东, 耿媛媛, 张贤明, 等, 2021. 产油微藻胁迫培养策略研究综述[J]. 中国环境科学, 41(8): 3853-3866.
|
| [8] |
王延莉, 曹阳, 梁祎凡, 等, 2022. 绵羊ACAA1基因多态性及其与肉质性状的关联性分析[J]. 吉林农业大学学报, 44(2): 204-209.
|
| [9] |
位文倩, 孙昕, 黄峰, 等, 2022. 转录组揭示自养和混合培养栅藻油脂代谢途径差异[J]. 中国油料作物学报, 44(1): 130-137.
|
| [10] |
于仁成, 吕颂辉, 齐雨藻, 等, 2020. 中国近海有害藻华研究现状与展望[J]. 海洋与湖沼, 51(4): 768-788.
|
| [11] |
张梅, 米铁柱, 甄毓, 等, 2018. 基于玛氏骨条藻转录组的脂肪酸合成途径分析[J]. 中国海洋大学学报, 48(4): 81-93.
|
| [12] |
周婧雯, 丁玉娇, 曾晓蓉, 等, 2020. 生菜种子低温处理后耐冻性和脂肪酸合成代谢的关系研究[J]. 亚热带植物科学, 49(1): 9-14.
|
| [13] |
庄珊珊, 2019. 海洋硅藻假微型海链藻休眠细胞形成过程的相关代谢调控[D]. 厦门: 厦门大学.
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}