
Journal of Tropical Oceanography >
Complete mitochondrial genome of Cladopsammia gracilis and Rhizopsammia wettsteini (Scleractinia, Dendrophylliidae) and its phylogenetic implications*
Copy editor: LIN Qiang
Received date: 2022-12-30
Revised date: 2023-03-14
Online published: 2023-03-21
Supported by
Guangdong Basic and Applied Basic Research Foundation(2022A1515010656)
Key Research and Development Project of Hainan Province(ZDYF2020200)
National Key Research and Development Program of China(2021YFF0502800)
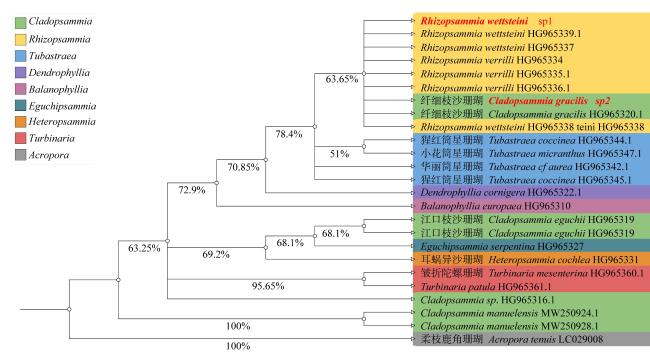
There is a taxonomic uncertainty on the species classification of Cladopsammia gracilis and Rhizopsammia wettsteini as well as the unresolved status of the closely related genera Cladopsammia and Rhizopsammia. In order to further analyze the phylogenetic relationship between C. gracilis and R. wettsteini, we examined the micromorphological and microstructural features and systematically compared the gene structure, genetic distance and codon preference of two mitochondrial genomes. The results show that there are three distinct characteristics: reptoid budding, costae and septa. The genome composition of the two species is very similar, containing 13 protein-coding genes, 2 rRNA genes and 2 tRNA genes. Unlike most scleractinians, there were no overlaps between genes. In addition, phylogenetic analysis based on CO1 genes show that C. gracilis and R. wettsteini are closely related but distantly related to other species of the genera Cladopsammia. The present study highlights the need for an integrative approach to conduct a deeper species delimitation of genera Cladopsammia and Rhizopsammia.
Key words: Dendrophylliidae; mitochondrial genomes; taxonomy; phylogenetic analysis
HUO Jiaxin , LI Yingxin , SONG Yan , ZHU Qing , ZHOU Weihua , YUAN Xiangcheng , HUANG Hui , LIU Sheng . Complete mitochondrial genome of Cladopsammia gracilis and Rhizopsammia wettsteini (Scleractinia, Dendrophylliidae) and its phylogenetic implications*[J]. Journal of Tropical Oceanography, 2024 , 43(3) : 22 -30 . DOI: 10.11978/2022267
表1 外源信息表Tab. 1 List of outer groups |
| 物种 | CO1基因序列 |
|---|---|
| 柔枝鹿角珊瑚Acropora tenuis | LC029008 |
| Balanophyllia europaea# | HG965310 |
| 江口枝沙珊瑚Cladopsammia eguchii | HG965319; HG965317.1 |
| 纤细枝沙珊瑚Cladopsammia gracilis | HG965320.1 |
| Cladopsammia sp.# | HG96531.6 |
| Cladopsammia mamuelensis# | MW250924.1; MW250928.1 |
| Dendrophyllia cornigera# | HG965322.1 |
| Eguchipsammia serpentina# | HG965327 |
| 耳蜗异沙珊瑚Heteropsammia cochlea | HG965331 |
| Rhizopsammia verrilli# | HG965336.1; HG965334; HG965335.1 |
| Rhizopsammia wettsteini# | HG965338; HG965337; HG965339.1 |
| 华丽筒星珊瑚Tubastraea cf. aurea | HG965342.1 |
| 猩红筒星珊瑚Tubastraea coccinea | HG965344 |
| 小花筒星珊瑚 Tubastraea micranthus | HG965347 |
| 皱折陀螺珊瑚Turbinaria mesenterina | HG965360 |
| Turbinaria patula# | HG965361 |
注: #表示该物种尚未有正式中文名 |
图1 Rhizopsammia wettsteini形态学特征a. 珊瑚群体形态与生殖根(reptoid budding, RB); b. 骨骼形态; c. 隔片排列情况; d. 轴柱; e. 珊瑚肋(costae, C)和肋间细沟(intercostal striae, IS) Fig. 1 The morphological characteristics of Rhizopsammia wettsteini. (a) Corallum and reptoid budding; (b) colony view; (c) septal arrangement; (d) dollumella, spongy; (e) costae (C) and intercostal striae (IS) porous |
图2 Cladopsammia gracilis形态学特征a. 珊瑚群体形态; b. 骨骼形态; c. 隔片排列情况; d. 轴柱; e. 珊瑚肋(costae, C)和肋间细沟(intercostal striae, IS) Fig. 2 The morphological characteristics of Cladopsammia gracilis. (a) Corallum; (b) colony view; (c) septal arrangement; (d) collumella, spongy; (e) costae (C) and intercostal striae (IS) porous |
表2 Rhizopsammia wettsteini和Cladopsammia gracilis线粒体基因组碱基组成Tab. 2 Composition of the Rhizopsammia wettsteini and Cladopsammia gracilis mitochondrial genome |
| 基因序列 | Rhizopsammia wettsteini | Cladopsammia gracilis | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| T/% | C/% | A /% | G /% | C+G /% | T/% | C/% | A /% | G /% | C+G /% | |
| 全基因组 | 39.7 | 16.0 | 21.1 | 23.2 | 39.4 | 37.4 | 13.6 | 25.4 | 23.7 | 37.2 |
| 蛋白编码基因 | 37.7 | 14.3 | 24.5 | 23.6 | 37.9 | 38.8 | 15.9 | 21.3 | 24.0 | 40.0 |
表3 Rhizopsammia wettsteini和Cladopsammia gracilis线粒体蛋白编码基因的相对同义密码子使用度(RSCU)Tab. 3 RSCU of Rhizopsammia wettsteini and Cladopsammia gracilis mitochondrial genome protein-coding sequence |
| 氨基酸 | 密码子 | 次数 | RSCU | 氨基酸 | 密码子 | 次数 | RSCU | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| R.wet* | C.gra | R.wet | C.gra | R.wet | C.gra | R.wet | C.gra | ||||
| Phe | UUU | 454 | 451 | 1.68 | 1.63 | Ser | UCU | 125 | 116 | 1.54 | 1.47 |
| UUC | 85 | 101 | 0.32 | 0.37 | UCC | 57 | 44 | 0.7 | 0.56 | ||
| Leu | UUA | 295 | 335 | 2.12 | 2.29 | UCA | 63 | 61 | 0.77 | 0.78 | |
| UUG | 207 | 216 | 1.49 | 1.48 | UCG | 50 | 55 | 0.61 | 0.7 | ||
| CUU | 162 | 165 | 1.16 | 1.13 | Pro | CCU | 52 | 62 | 1.25 | 1.22 | |
| CUC | 49 | 48 | 0.35 | 0.33 | CCC | 53 | 51 | 1.28 | 1.00 | ||
| CUA | 69 | 65 | 0.5 | 0.44 | CCA | 31 | 43 | 0.75 | 0.85 | ||
| CUG | 54 | 48 | 0.39 | 0.33 | CCG | 30 | 47 | 0.72 | 0.93 | ||
| Ile | AUU | 262 | 240 | 1.7 | 1.68 | Thr | ACU | 82 | 76 | 1.63 | 1.70 |
| AUC | 56 | 64 | 0.36 | 0.45 | ACC | 41 | 29 | 0.82 | 0.65 | ||
| AUA | 143 | 124 | 0.93 | 0.87 | ACA | 55 | 46 | 1.09 | 1.03 | ||
| Met | AUG | 121 | 131 | 1 | 1.00 | ACG | 23 | 28 | 0.46 | 0.63 | |
| Val | GUU | 233 | 203 | 1.93 | 1.75 | Ala | GCU | 110 | 101 | 1.62 | 1.60 |
| GUC | 58 | 77 | 0.48 | 0.67 | GCC | 60 | 53 | 0.88 | 0.84 | ||
| GUA | 83 | 86 | 0.69 | 0.74 | GCA | 42 | 51 | 0.62 | 0.81 | ||
| GUG | 110 | 97 | 0.91 | 0.84 | GCG | 60 | 48 | 0.88 | 0.76 | ||
| Tyr | UAU | 197 | 192 | 1.66 | 1.54 | Cys | UGU | 136 | 130 | 1.42 | 1.38 |
| UAC | 41 | 57 | 0.34 | 0.46 | UGC | 56 | 59 | 0.58 | 0.62 | ||
| Asp | GAU | 93 | 88 | 1.45 | 1.30 | Glu | GAA | 111 | 118 | 1.06 | 1.11 |
| GAC | 35 | 47 | 0.55 | 0.70 | Trp | UGG | 160 | 168 | 1 | 1.00 | |
| His | CAU | 68 | 54 | 1.49 | 1.35 | Arg | CGU | 30 | 23 | 0.58 | 0.42 |
| CAC | 23 | 26 | 0.51 | 0.65 | CGC | 20 | 14 | 0.38 | 0.26 | ||
| Gln | CAA | 85 | 65 | 1.4 | 1.30 | CGA | 44 | 41 | 0.85 | 0.75 | |
| CAG | 36 | 35 | 0.6 | 0.70 | CGG | 35 | 59 | 0.67 | 1.09 | ||
| Asn | AAU | 165 | 150 | 1.47 | 1.47 | AGA | 119 | 95 | 2.29 | 1.75 | |
| AAC | 59 | 54 | 0.53 | 0.53 | AGG | 64 | 94 | 1.23 | 1.73 | ||
| Lys | AAA | 179 | 186 | 1.18 | 1.23 | Ser | AGU | 148 | 149 | 1.82 | 1.89 |
| AAG | 124 | 116 | 0.82 | 0.77 | AGC | 45 | 47 | 0.55 | 0.60 | ||
| TER | UAA | 119 | 167 | 1.17 | 1.43 | Gly | GGU | 137 | 139 | 1.09 | 1.15 |
| UAG | 93 | 87 | 0.91 | 0.75 | GGC | 85 | 69 | 0.67 | 0.57 | ||
| UGA | 93 | 96 | 0.91 | 0.82 | GGA | 92 | 92 | 0.73 | 0.76 | ||
注: * R.wet和C.gra分别代表Rhizopsammia wettsteini和Cladopsammia gracilis |
表4 Rhizopsammia wettsteini和Cladopsammia gracilis线粒体基因组密码子偏好性主要参数Tab. 4 Usage bias parameters of Rhizopsammia wettsteini and Cladopsammia gracilis mitochondrial genome |
| 密码子偏好性主要参数 | R. wettsteini | C. gracilis |
|---|---|---|
| T3s | 0.477 | 0.459 |
| C3s | 0.160 | 0.164 |
| A3s | 0.323 | 0.327 |
| G3s | 0.277 | 0.289 |
| CAI | 0.148 | 0.143 |
| CBI | -0.152 | -0.151 |
| Fop | 0.310 | 0.307 |
| ENc | 49.90 | 51.07 |
| CG3 | 0.330 | 0.343 |
| GC | 0.381 | 0.384 |
| [1] |
戴昌凤, 洪圣雯, 2009. 台湾珊瑚图鉴[M]. 台北: 猫头鹰出版社: 1-256 (in Chinese).
|
| [2] |
戴昌凤, 郑有容, 2020. 台湾珊瑚全图鉴(上): 石珊瑚[M]. 台北: 猫头鹰出版社: 1-557 (in Chinese).
|
| [3] |
倪智, 刘佳英, 宋倩倩, 等, 2017. 厦门白哈礁海域石珊瑚的分类鉴定[J]. 应用海洋学学报, 36(4): 550-557.
|
| [4] |
张喆, 俞晓磊, 梁宇娴, 等, 2022. 海水升温对非虫黄藻共生型珊瑚Cladopsammia sp. 的生理影响[J]. 热带海洋学报, 41(5): 141-149.
|
| [5] |
赵东波, 杨顺良, 姬厚德, 2022. 厦门同安湾海域猩红筒星珊瑚移植初步试验[J]. 渔业研究, 44(3): 275-283.
|
| [6] |
邹仁林, 2001. 中国动物志, 腔肠动物门, 珊瑚虫纲, 石珊瑚目, 造礁石珊瑚[M]. 北京: 科学出版社: 1-289.
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
/
| 〈 |
|
〉 |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}