
Journal of Tropical Oceanography >
Copulation, embryonic and post-embryonic development of Sphaeramia nematoptera
Copy editor: SUN Cuici
Received date: 2023-03-24
Revised date: 2023-04-18
Online published: 2023-05-25
Supported by
Guided Project of Fujian Provincial Department of Science and Technology(2022N0014)
The Regional Demonstration Project of the 13th Five-Year Plan of Marine Economy Innovation & Development in Xiamen(16PZY002SF18)
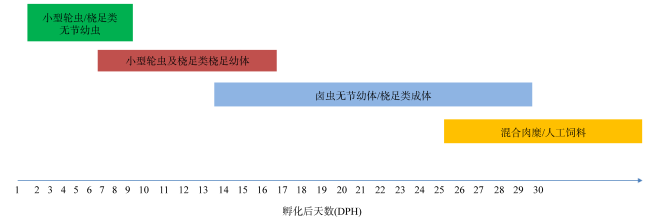
The copulation behavior, embryonic and post-embryonic development of Sphaeramia nematoptera were investigated in this study. The results showed that the mating behavior was dominated by females, mating between 1:00 and 3:00 am, and the reproductive cycle was 11~18 d without seasonal fluctuations. The release of eggs by female were 3126~4882 at a time, which were incubated by the male’s mouth. The fertilized eggs were suborbicular, (720±10) μm in diameter, with several oil globules inside. It took 187 h for a fertilized egg to be hatched under (27.0±0.5) ℃. The embryo development process included Cleavage, Blastocyst, Gastrulation, Neurula and Organ formation stages. The post-embryonic development could be divided into juvenile stage, larval stage, juvenile stage and adult stage. The whole length of the first hatchling was (2.8±0.1) mm, the height of the mouth cleft was (0.202±18) mm, and the juvenile opened its mouth to feed at the second day after hatching. Fins began to form and two black bands appeared on the body when ending the floating stage and entering the larval stage 24d after hatching. Scales were fully formed 50d after hatching and the body color and markings as well as other appearance characteristics were similar to the adult fish, which indicated that the fish entered the juvenile stage. Maturation of gonad was achieved 360d after hatching.
LAN Zhenqiang , ZHENG Jitao , CHEN Yun , CHEN Nan , WANG Shuhong . Copulation, embryonic and post-embryonic development of Sphaeramia nematoptera[J]. Journal of Tropical Oceanography, 2024 , 43(1) : 116 -125 . DOI: 10.11978/2023039
图2 丝鳍圆天竺鲷的交配行为及含卵过程a. 雄鱼(左)鼓起下颌回应雌鱼; b. 雄鱼(右)和雌鱼在缸的一角并排游动; c. 雌鱼(左)产下卵团(如箭头所示); d. 雄鱼(右)转身将卵团含入口中(如箭头所示) Fig. 2 The mating behavior and oral incubation of S. nematoptera. (a) Male (left) puffs out jaws in response to female; (b) the male (right) and the female are swimming side by side in one corner of the tank (as shown by the arrow); (c) female (left) laying egg mass; (d) male (right) turns around and takes the egg mass into mouth (as shown by the arrow) |
表1 丝鳍圆天竺鲷在2021年3月—5月产卵情况Tab. 1 Spawning of S. nematoptera during March to May 2021 |
| 亲鱼组 | 序号 | 产卵日期 /(月.日) | 产卵时间 /(时: 分) | 孵化用时/h | 产卵间隔/d |
|---|---|---|---|---|---|
| A | A-1 | 03.08 | 01:31 | 187 | / |
| A-2 | 03.19 | 02:12 | 187.5 | 11 | |
| A-3 | 04.05 | 01:40 | 187 | 17 | |
| B | B-1 | 03.09 | 02:01 | 187.5 | / |
| B-2 | 03.24 | 01:39 | 187 | 15 | |
| B-3 | 04.11 | 01:22 | 187 | 18 | |
| C | C-1 | 03.01 | 01:09 | 186.5 | / |
| C-2 | 03.14 | 01:33 | 187 | 13 | |
| C-3 | 03.29 | 01:29 | 187 | 15 | |
| D | D-1 | 03.27 | 02:18 | 188 | / |
| D-2 | 04.13 | 01:56 | 187.5 | 17 | |
| D-3 | 05.01 | 01:57 | 187.5 | 18 |
注: 表格中/为2021年3—5月记录中的该亲鱼组的第一次产卵 |
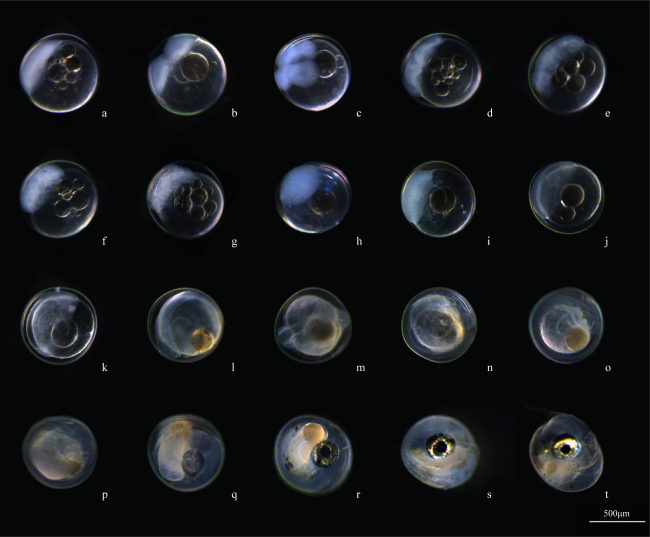
表2 丝鳍圆天竺鲷胚胎发育时序Tab. 2 Embryonic development of S. nematoptera |
| 发育时期 | 受精后时间 | 各发育期简要特征 | 图版 |
|---|---|---|---|
| 胚盘隆起期 | 30min | 呈圆形、半透明, 卵之间有黏丝相连 | 4a |
| 2细胞期 | 1h | 2个分裂球 | 4b |
| 4细胞期 | 1h 30min | 继续分裂为4个分裂球 | 4c |
| 8细胞期 | 2h 30min | 再次分裂, 此时有8个分裂球 | 4d |
| 16细胞期 | 3h 10min | 分裂为16个分裂球 | 4e |
| 32细胞期 | 4h | 32个分裂球 | 4f |
| 64细胞期 | 5h 15min | 分裂球很小, 界限不明, 有64个分裂球 | 4g |
| 囊胚早期 | 6h 40min | 看不清细胞界限, 分裂球组成的囊胚高举 | 4h |
| 囊胚中期 | 9h 15min | 举起的细胞变扁平, 囊胚层更低 | 4i |
| 囊胚晚期 | 12h 45min | 囊胚层变扁, 并向卵黄部分下包 | 4j |
| 原肠期 | 22h 30min | 胚盘下包, 胚环和胚盾出现 | 4k |
| 器官形成期(眼囊期) | 29h 15min | 油滴变小, 出现圆形的眼囊 | 4l |
| 器官形成期(尾芽期) | 34h | 体节10~12对, 胚体后端出现尾芽 | 4m |
| 器官形成期 | 56h 30min | 器官继续发育, 可见嗅囊、耳囊 | 4n |
| 器官形成期 | 67h | 脑径增大, 体节增多心脏开始搏动 | 4o |
| 器官形成期 | 80h 30min | 眼圈出现黑色素, 卵黄变小 | 4p |
| 器官形成期 | 112h | 心脏跳动加快, 身体上出现黑色素 | 4q |
| 器官形成期 | 136h | 眼部积聚鸟粪素, 可见明显口裂 | 4r |
| 孵化前期 | 160h | 胚体在膜内转动, 血液循环明显 | 4s |
| 孵化期 | 187h | 身体破膜 | 4t |
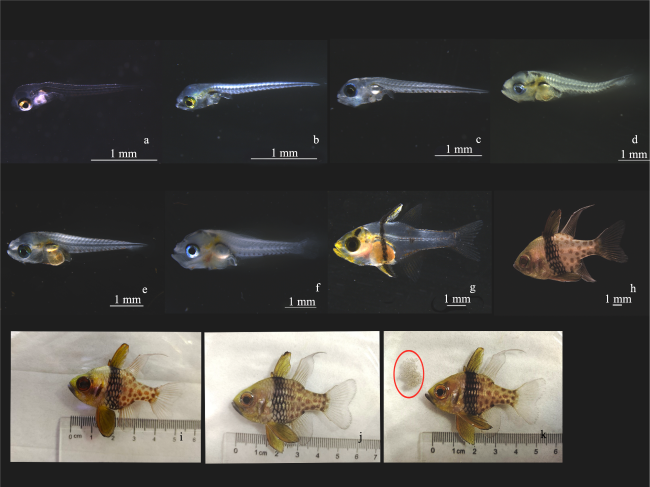
图5 丝鳍圆天竺鲷各个发育阶段的显微图片和照片a. 初孵仔鱼, 全长(2.82±0.12)mm, 趋光性强; b. 1 DPH仔鱼, 身体透明, 可开口摄食; c. 2 DPH仔鱼全长(3.54±0.23)mm, 卵黄囊大部分被消耗; d. 4 DPH仔鱼口裂高度(0.23±0.02)mm, 消化道内可见大量饵料; e. 12 DPH仔鱼全长(4.42±0.38)mm, 腹部和头部色素沉积明显; f. 18 DPH仔鱼身体变宽, 背鳍和臀鳍鳍条出现; g. 24 DPH稚鱼, 体型近似成鱼, 背鳍分为2个, 鳞片开始出现; h. 50 DPH幼鱼, 鳞片完全形成, 体型与体表斑纹与成鱼相同; i. 200 DPH幼鱼, 性腺未成熟; j. 360 DPH性成熟雌鱼, 全长(53.13±2.11)mm, 性腺达到初次性成熟, 雌性个体腹部较为膨大; k. 360 DPH性成熟雄鱼, 红圈内为吐出的受精卵 Fig.5 Photomicrographs and photographs of the various stages of development of S. nematopter. (a) Newly hatched larvae were (2.82±0.12) mm in total length, with strong phototropism; (b) 1 DPH larvae with transparent bodies ready for feeding; (c) 2 DPH larvae were (3.54±0.23) mm in total length, and most of the yolk-sac was consumed; (d) 4 DPH larvae with a mouth cleft height of (0.23±0.02) mm and a large amount of prey visible in the stomach; (e) 12 DPH larvae were (4.42±0.38) mm in total length, with marked pigmentation on the abdomen and head; (f) 18 DPH larvae with broadened bodies and the presence of dorsal and anal fin flippers; (g) 24 DPH juvenile with morphology approximating that of adult, with the dorsal fin divided into 2 and scales beginning to appear; (h) 50 DPH juvenile with fully formed scales and the same body shape and body markings as the adult; (i) 200 DPH young fish with immature gonads; (j) 360 DPH sexually mature female, (53.13±2.11) mm in total length, gonads reaching first sexual maturity, females with more expanded abdomens; (k) 360DPH sexually mature male, with spatulated fertilized eggs in red circle |
表3 丝鳍圆天竺鲷与其他天竺鲷受精卵和初孵仔鱼特性比较Tab. 3 Comparison of the characteristics of fertilized eggs and first hatchlings of S. nematoptera and other cardinalfish |
| 物种 | 卵径大小/mm | 胚胎发育时间/d | 初孵仔鱼长度/mm | 文献来源 |
|---|---|---|---|---|
| 四眼天竺鲷(Apogon quadrifasciatus) | 0.45 | 5 | 2.6 | Saravanan et al (2013) |
| 欧洲天竺鲷(A. imberbis) | 0.5 | 5~7 | 2 | Mazzoldi et al (2008) |
| 犬牙天竺鲷(A. affinis) | 0.35~0.4 | / | 1 | Smith et al (1971) |
| 半线天竺鲷(A. semilineatus) | 0.58~0.6 | / | 2.3 | Ebina (1932) |
| 拉氏天竺鲷(A. rueppellii) | 2.4 | 16 | 5.5~6.4 | Chrystal et al (1985) |
| 考氏鳍竺鲷(P. kauderni) | 2.8~3.2 | 19 | 6~6.5 | Vagelli (1999) |
| 环纹圆天竺鲷(S. orbicularis) | 0.6~0.7 | 8 | 3.3 | Allen (1975) |
| 丝鳍圆天竺鲷(S. nematoptera) | 0.7 | 8 | 2.8 | 本文章 |
| [1] |
王军, 陈明茹, 谢仰杰, 2008. 鱼类学[M]. 厦门: 厦门大学出版社: 240-241.
|
| [2] |
殷名称, 1991. 鱼类早期生活史研究与其进展[J]. 水产学报, 15(4): 348-358.
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}