
Journal of Tropical Oceanography >
Influence of tree species and intertidal elevations on the carbon storage of the Gaoqiao mangrove area in Zhanjiang, Guangdong Province
Copy editor: YIN Bo
Received date: 2023-06-05
Revised date: 2023-08-28
Online published: 2023-08-30
Supported by
Special Funds for Natural Resource Ecological Restoration Projects in Guangdong Province(440000210000000019650)
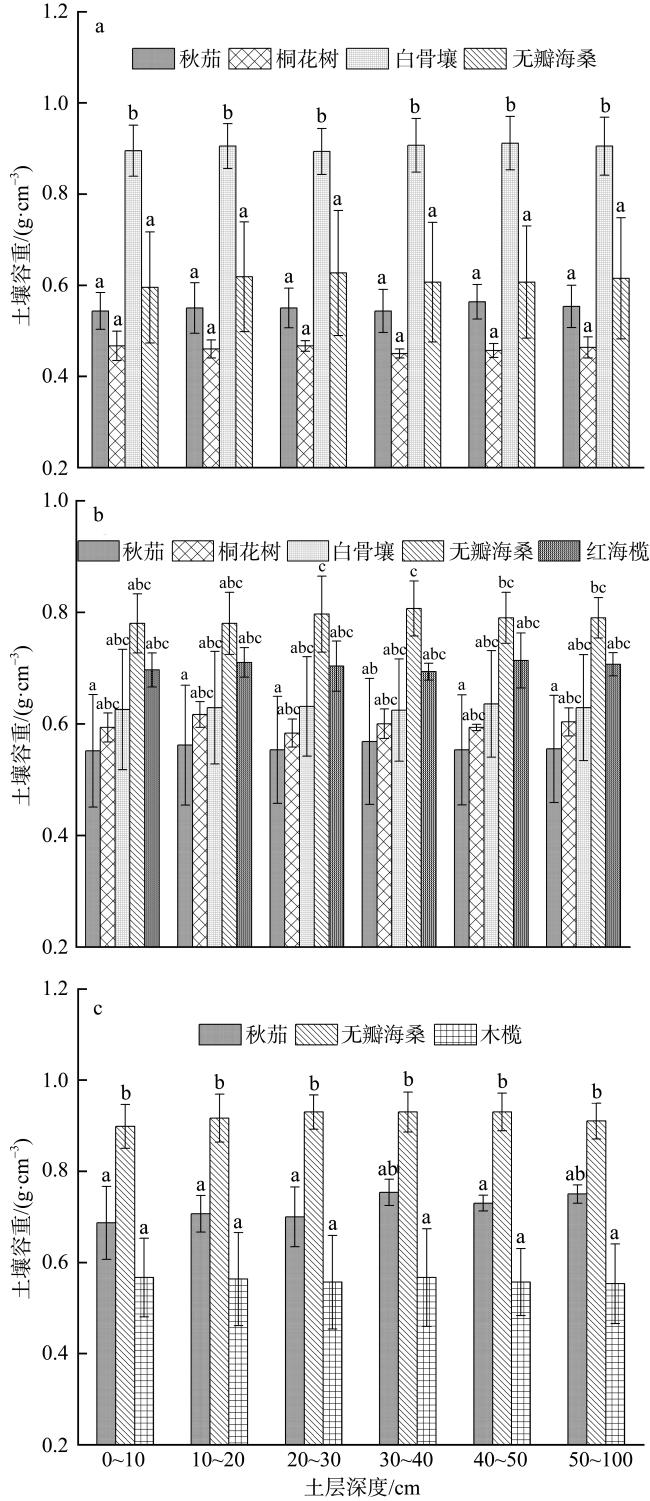
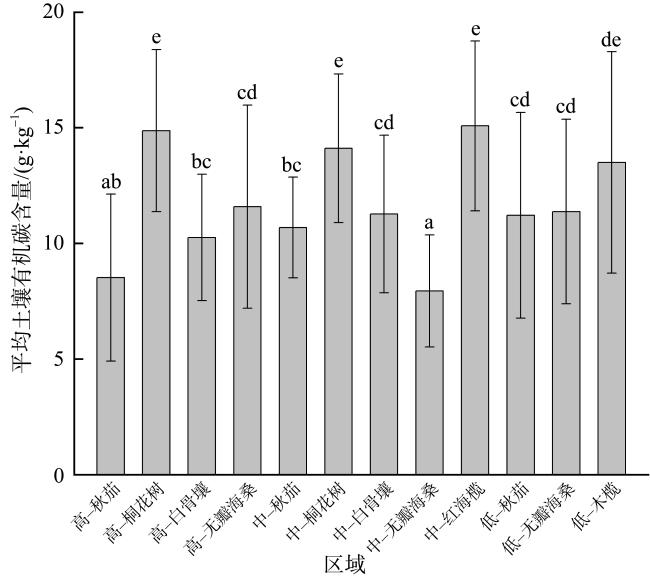
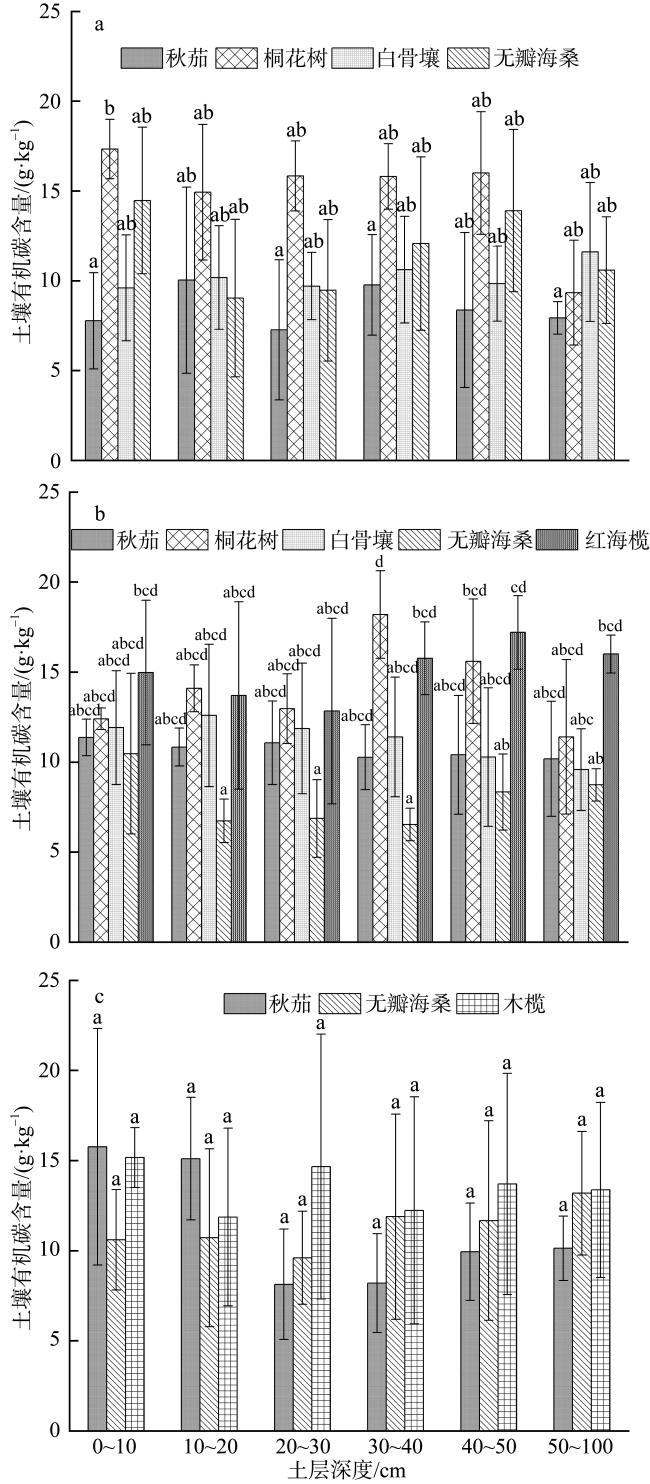
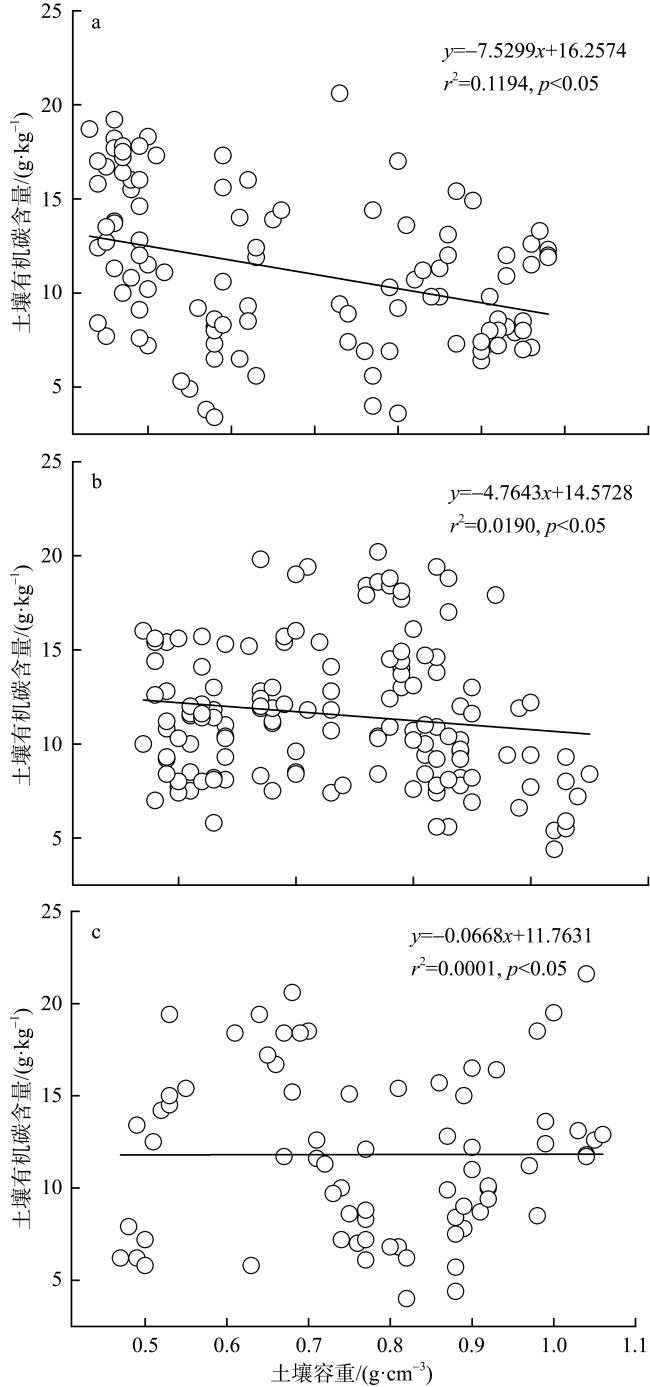
Mangroves are located at the junction between marine and land, and their carbon storage capacity depends on a variety of factors. In this study, the plant growth, biomass, soil organic carbon content and soil bulk density as well as the carbon storage in both vegetation and soil were compared in the mangrove communities in the Gaoqiao District, Zhanjiang, Guangdong, to study the individual and coupled effects of tree species and intertidal elevation factors on carbon storage. The results showed that carbon storage of mangrove communities was 4.98×1010 g and carbon density was 107.76×106 g·hm-2, and the carbon storage was mainly concentrated in soil. The carbon density of mangrove in different intertidal elevations were significantly affected by tree species, and the carbon storages of mangrove at low intertidal elevation was higher than those at middle and high intertidal elevation. The carbon storage of Sonneratia apetala communities was significantly affected by intertidal elevation. The carbon storage of Rhizophora stylosa communities at middle intertidal elevation and S. apetala communities at low intertidal elevation was greatest in this mangrove area, regardless of the perspective of vegetation carbon pool or soil carbon pool. In all mangrove communities, the vertical distribution of soil organic carbon content was homogeneous with no significant difference among soil layers. The vertical distribution of soil organic carbon contents at the high and middle intertidal elevation was influenced by tree species. Among them, the soil organic carbon content in the majority of Aegiceras corniculatum communities was significantly higher than those at the same soil layer of Kandelia obovata, Avicennia marina and S. apetala communities. However, tree species had no significant effect on the soil organic carbon content at the low intertidal elevation. Although the relationship of soil bulk density and soil organic carbon content was influenced by both tree species and intertidal elevation, the linear fitting results were mainly negative correlations. This study can provide a scientific basis for the selection of tree species and intertidal elevation for carbon sequestration in mangrove afforestation project.
ZHOU Zhigang , YUE Wen , LI Huiquan , LIN Yangyang . Influence of tree species and intertidal elevations on the carbon storage of the Gaoqiao mangrove area in Zhanjiang, Guangdong Province[J]. Journal of Tropical Oceanography, 2024 , 43(2) : 108 -120 . DOI: 10.11978/2023074
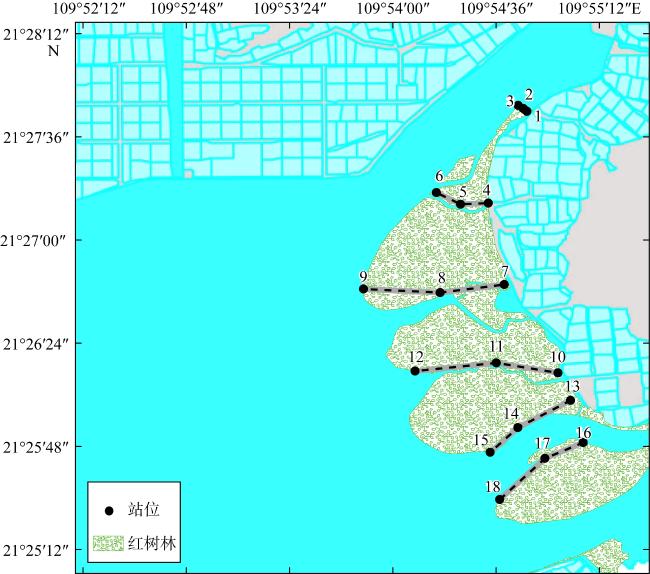
图1 高桥红树林采样站位分布示意图数字表示站位名称, 数字间的灰色和虚线表示断面。该图基于自然资源部标准地图服务站下载的审图号粤S(2018)099号标准地图制作 Fig. 1 Distribution of sampling sites in the Gaoqiao mangrove area |
表1 红树林采样站位基本信息Tab. 1 The general features of sampling sites in the Gaoqiao mangrove area |
| 站位 | 红树群落类型 | 潮滩高程 |
|---|---|---|
| 1 | 秋茄 | 中潮滩 |
| 2 | 秋茄 | 高潮滩 |
| 3 | 桐花树 | 高潮滩 |
| 4 | 无瓣海桑 | 高潮滩 |
| 5 | 白骨壤 | 中潮滩 |
| 6 | 无瓣海桑 | 中潮滩 |
| 7 | 秋茄 | 中潮滩 |
| 8 | 白骨壤 | 高潮滩 |
| 9 | 白骨壤 | 中潮滩 |
| 10 | 木榄 | 低潮滩 |
| 11 | 白骨壤 | 中潮滩 |
| 12 | 无瓣海桑 | 高潮滩 |
| 13 | 红海榄 | 中潮滩 |
| 14 | 白骨壤 | 中潮滩 |
| 15 | 白骨壤 | 高潮滩 |
| 16 | 无瓣海桑 | 低潮滩 |
| 17 | 秋茄 | 低潮滩 |
| 18 | 无瓣海桑 | 低潮滩 |
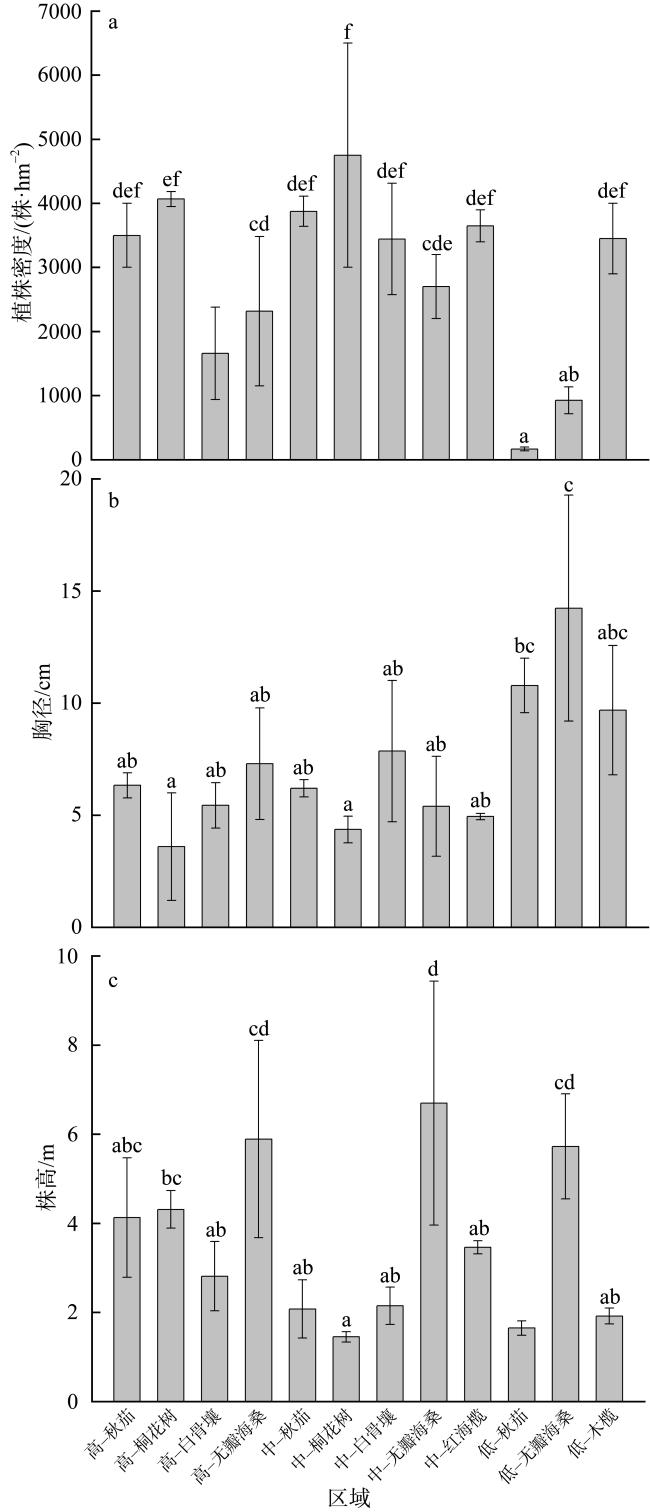
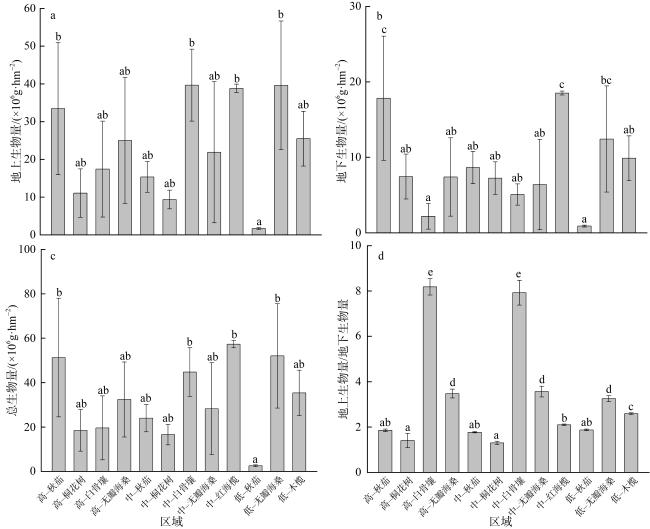
表2 不同红树林树种的地上及地下生物量模型Tab. 2 Aboveground and underground biomass allometric equations of different tree species |
| 树种 | 地上生物量模型 | 地下生物量模型 | 参考文献 |
|---|---|---|---|
| 秋茄(Kandelia obovata) | Ba=0.0341(d2h)1.03 | Bu=0.0483(d2h)0.834 | Hoque等(2011) |
| 无瓣海桑(Sonneratia apetala) | Ba=0.280(d2h)0.693 | Bu=0.038(d2h)0.759 | Ren等(2005) |
| 桐花树(Aegiceras corniculatum) | lgBa=1.496+0.465lg(d2h) | lgBu=0.967+0.303lg(d2h) | Tam等(1995) |
| 白骨壤(Avicennia marina) | lgBa=2.113lgd-0.511 | lgBu=1.171lgd+0.106 | Comley等(2005) |
| 木榄(Bruguiera gymnorrhiza) | Ba=0.168d2.31 | Bu=0.0188(d2h)0.909 | Clough等(1989); Tam等(1995) |
| 红海榄(Rhizophora stylosa) | Ba=0.2206d2.4292 | Bu=0.261d1.86 | Tamai等(1988); Fromard等(1998) |
注: Ba为地上生物量(单位: ×106g); Bu为地下生物量(单位: ×106g); d为胸径(单位: cm); h为树高(单位: m) |
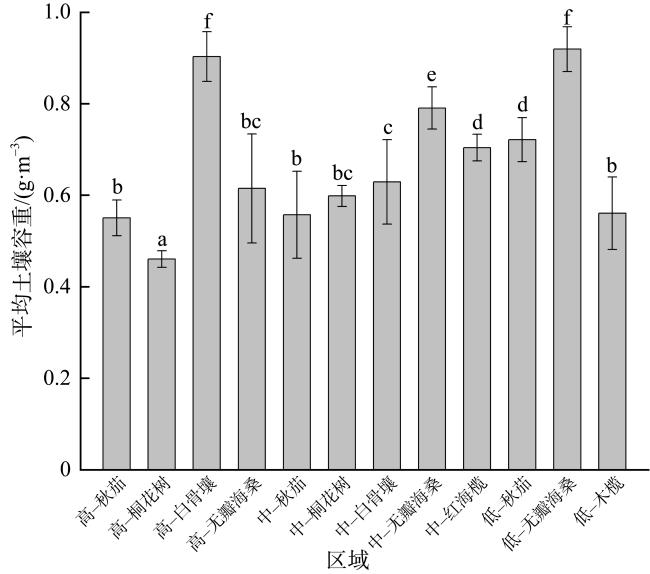
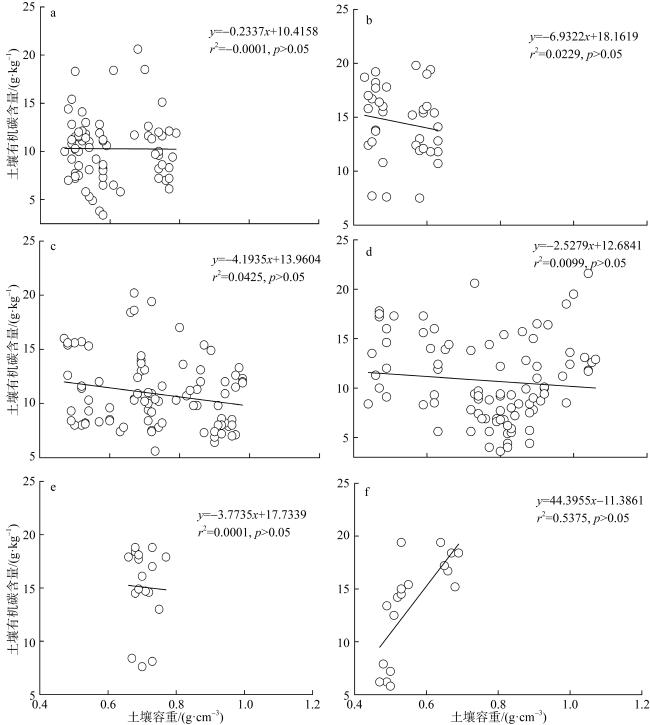
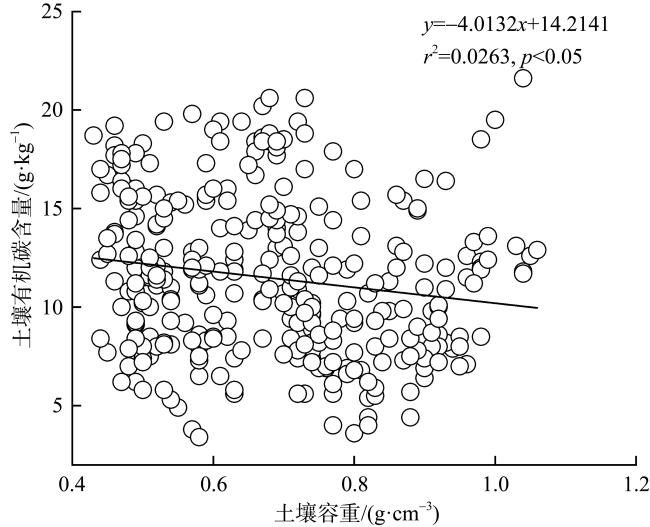
图8 不同潮滩高程的土壤有机碳含量与土壤容重的相关性分析a. 高潮滩; b. 中潮滩; c. 低潮滩 Fig. 8 Correlation analysis between soil organic carbon content and soil bulk density at different intertidal elevations |
图9 不同红树群落的土壤有机碳含量与土壤容重的线性回归关系a. 秋茄; b. 桐花树; c. 白骨壤; d. 无瓣海桑; e. 红海榄; f. 木榄 Fig. 9 Linear regression relationship between soil organic carbon content and soil bulk density in different mangrove communities |
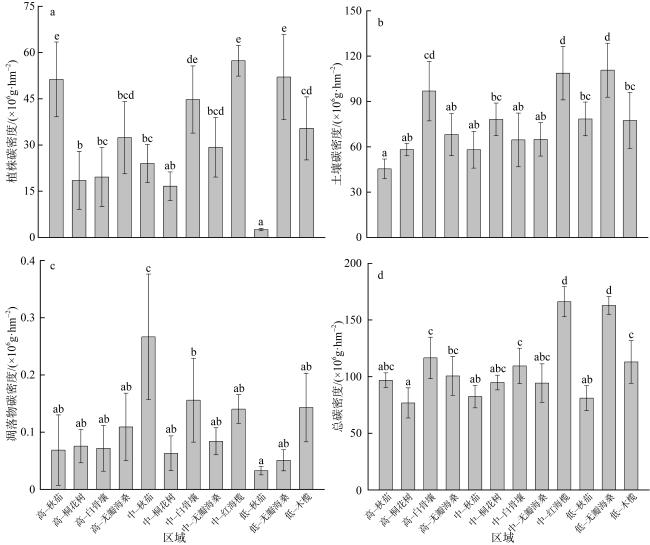
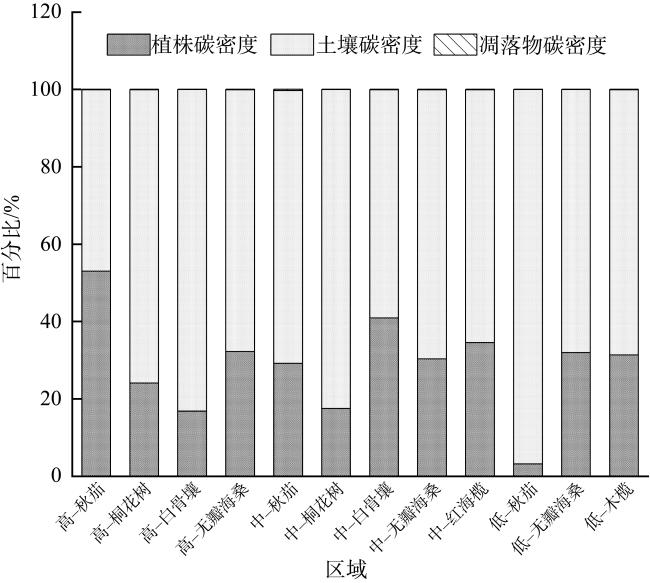
图11 不同红树群落的碳密度分布不同小写字母表示差异显著(p<0.05) Fig. 11 Carbon density distribution of different mangrove communities (different lowercase letters show significant difference, p< 0.05) |
| [1] |
曹庆先, 2010. 北部湾沿海红树林生物量和碳贮量的遥感估算[D]. 北京: 中国林业科学研究院:99-104.
|
| [2] |
陈鹭真, 林鹏, 王文卿, 2006. 红树植物淹水胁迫响应研究进展[J]. 生态学报, 26(2): 586-593.
|
| [3] |
陈玮琪, 洪华生, 张珞平, 等, 2007. 中国红树林生态系统服务的能值价值[J]. 资源科学, 29(1): 147-154.
|
| [4] |
陈小花, 陈宗铸, 雷金睿, 等, 2022. 清澜港红树林湿地典型群落类型沉积物活性有机碳组分分布特征[J]. 生态学报, 42(11): 4572-4581.
|
| [5] |
陈瑶瑶, 张雅松, 娄铎, 等, 2019. 广东英罗湾不同潮位红树林-滩涂系统碳密度差异[J]. 生态环境学报, 28(6): 1134-1140.
|
| [6] |
陈玉军, 李婷, 朱立安, 等, 2022. 淹水梯度对红树林湿地土壤微生物生物量与酶活性的影响[J]. 水土保持通报, 42(6): 68-75.
|
| [7] |
成家隆, 吕瑜良, 陈玉军, 等, 2015. 水东湾红树林不同生长位置生长差异性研究[J]. 生态科学, 34(6): 30-35.
|
| [8] |
高天伦, 管伟, 毛静, 等, 2017. 广东省雷州附城主要红树林群落碳储量及其影响因子[J]. 生态环境学报, 26(6): 985-990.
|
| [9] |
胡倩芳, 2009. 不同高程下白骨壤+秋茄人工红树林生长、生理及其更新能力的比较研究[D]. 厦门: 厦门大学:15-32.
|
| [10] |
胡懿凯, 2019. 淇澳岛不同恢复类型红树林碳密度和固碳速率比较研究[D]. 长沙: 中南林业科技大学:17-23.
|
| [11] |
黄诚智, 曾德华, 2019. 4种红树树种在三亚河红树林自然保护区的生长和生理生化响应[J]. 热带林业, 47(1): 34-37.
|
| [12] |
黄灵玉, 2015. 广东红树林土壤有机碳分布特征及其影响因素研究[D]. 桂林: 广西师范学院.
|
| [13] |
黄星, 袁菁菁, 马基仪, 等, 2022. 沙井港不同湿地植被群落沉积物中有机碳含量与分布特征[J]. 科技技术创新, (2): 25-28.
|
| [14] |
来洪运, 王咏雪, 章翊涵, 等, 2021. 浙江苍南沿浦湾秋茄人工林早期生长特征研究[J]. 林业科学研究, 34(4): 156-165.
|
| [15] |
李林锋, 吴小凤, 刘素青, 2015. 湛江5种红树林树种光合作用特性及光合固碳能力研究[J]. 广西植物, 35(6): 825-832.
|
| [16] |
李燕, 赵志忠, 王鸿平, 等, 2018. 海南东寨港红树林湿地沉积物有机碳的分布特征[J]. 安徽农业大学学报, 45(2): 268-273.
|
| [17] |
廖宝文, 2010. 三种红树植物对潮水淹浸与水体盐度适应能力的研究[D]. 北京: 中国林业科学研究院:17-41.
|
| [18] |
罗美娟, 2012. 红树植物桐花树幼苗对潮汐淹水胁迫的响应研究[D]. 北京: 中国林业科学研究院:15-23.
|
| [19] |
孙学超, 黄展鹏, 张琼锐, 等, 2022. 珠海淇澳岛人工次生无瓣海桑纯林的植被碳储量变化[J]. 华南师范大学学报(自然科学版), 54(4): 89-100.
|
| [20] |
辛琨, 颜葵, 李真, 等, 2014. 海南岛红树林湿地土壤有机碳分布规律及影响因素研究[J]. 土壤学报, 51(5): 1078-1086.
|
| [21] |
许大全, 张玉忠, 张荣铣, 1992. 植物光合作用的光抑制[J]. 植物生理学通讯, 28(4): 237-243.
|
| [22] |
张莉, 2013. 海南清澜港红树林土壤有机碳及其与土壤因子关系研究[D]. 洛阳: 河南科技大学:29-30.
|
| [23] |
张莉, 郭志华, 李志勇, 2013. 红树林湿地碳储量及碳汇研究进展[J]. 应用生态学报, 24(4): 1153-1159.
|
| [24] |
郑鹏, 史红文, 邓红兵, 等, 2012. 武汉市65个园林树种的生态功能研究[J]. 植物科学学报, 30(5): 468-475.
|
| [25] |
周莉, 李保国, 周广胜, 2005. 土壤有机碳的主导影响因子及其研究进展[J]. 地球科学进展, 20(1): 99-105.
|
| [26] |
朱耀军, 赵峰, 郭菊兰, 等, 2017. 广东湛江高桥红树林湿地沉积柱粒度特征[J]. 北京林业大学学报, 39(11): 9-17.
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}