
Journal of Tropical Oceanography >
Community structure of scleractinian corals in the northern South China Sea and their responses to the marine heatwaves
Copy editor: LIN Qiang
Received date: 2023-08-13
Revised date: 2023-10-20
Online published: 2023-10-31
Supported by
National Natural Science Foundation of China(42277362)
National Natural Science Foundation of China(41806139)
Science and Technology Planning Project of Guangzhou City(202002030345)
Central Public-Interest Scientific Institution Basal Research Fund(PM-zx703-202105-176)
Central Public-Interest Scientific Institution Basal Research Fund(PM-zx703-202004-143)
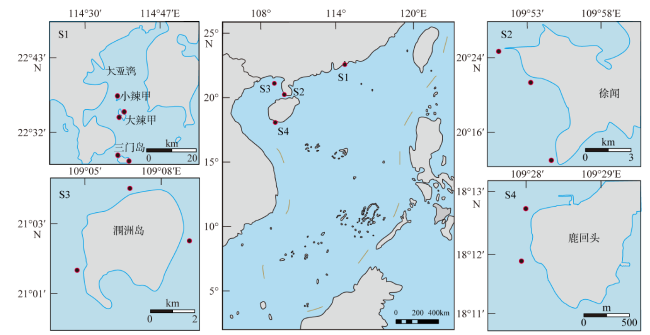
Coral communities in the northern South China Sea (SCS), which have the potential to act as regions of refugia for many coral species in the face of global warming. However, the temporal and spatial diversities of coral communities in the northern SCS and the effects of coral bleaching caused by marine heat waves are still unclear. In this study, the status of coral communities across the northern SCS, including the Daya Bay, the Weizhou Island, the Xuwen and Luhuitou fringing reefs, and the space differences are studied on the basis of ecological materials in the field investigations during August 2020, and its response to marine heatwaves. Our results indicate that coral communities across the northern SCS show remarkable regional characteristics. The dominant species composition, species number and α diversity index of coral communities are subject to significant regional differences, and the live coral cover is higher in the Luhuitou and Daya Bay (37.78% and 17.97%, respectively). Thus, the coral communities in Xuwen and Weizhou were subject to mass coral bleaching events during the field investigation, and the coral bleaching rate and mortality rate was 84.78% and 10.68%, 65.52% and 0.70% in Xuwen and the Weizhou Island, respectively, while no coral bleaching occurred in the Daya Bay and Luhuitou. Based on historical data analysis, although the live coral cover of coral communities in the northern SCS has dropped significantly since 1980s, there has been a recovery trend in recent years; the proportion of heat bleaching tolerant genera and heat bleaching resistant genera increased in the coral communities of the Weizhou Island, Xuwen and Luhuitou. In general, the coral community in the northern SCS has a strong resistance to heat bleaching. Strengthening the conservation and management of coral reef areas is crucial for the restoration and refuge of coral communities in the northern SCS under global warming.
XU Lijia , LIAO Zhiheng , CHEN Hui , WANG Yongzhi , HUANG Baiqiang , LIN Qiaoyun , GAN Jianfeng , YANG Jing . Community structure of scleractinian corals in the northern South China Sea and their responses to the marine heatwaves[J]. Journal of Tropical Oceanography, 2024 , 43(3) : 58 -71 . DOI: 10.11978/2023118
附表1 南海北部沿岸海域造礁石珊瑚种类(2020年)Appendix 1 List of scleractinian coral species on coral reefs of the northern South China Sea in 2020 |
| 类群 | 科 | 属 | 物种 | 珊瑚礁区 | 耐热性类型 | |||
|---|---|---|---|---|---|---|---|---|
| 大亚湾 | 涠洲岛 | 徐闻 | 鹿回头 | |||||
| 复 杂 类 群 | 鹿角珊瑚科 Acroporidae | 鹿角珊瑚属 Acropora | 粗野鹿角珊瑚Acropora humilis | + | + | 热敏感型 | ||
| 风信子鹿角珊瑚 Acropora hyacinthus | + | 热敏感型 | ||||||
| 彰显鹿角珊瑚Acropora insignis | + | 热敏感型 | ||||||
| 美丽鹿角珊瑚Acropora muricata | + | 热敏感型 | ||||||
| 多孔鹿角珊瑚Acropora millepora | + | 热敏感型 | ||||||
| 霜鹿角珊瑚Acropora pruinosa | + | + | 热敏感型 | |||||
| 单独鹿角珊瑚Acropora solitaryensis | + | 热敏感型 | ||||||
| 柔枝鹿角珊瑚Acropora tenuis | + | 热敏感型 | ||||||
| 强壮鹿角珊瑚Acropora valida | + | 热敏感型 | ||||||
| 蔷薇珊瑚属 Montipora | 繁锦蔷薇珊瑚 Montipora efflorescens | + | + | 热敏感型 | ||||
| 贺氏蔷薇珊瑚 Montipora hoffmeisteri | + | 热敏感型 | ||||||
| 变形蔷薇珊瑚 Montipora informis | + | 热敏感型 | ||||||
| 翼形蔷薇珊瑚 Montipora peltiformis | + | 热敏感型 | ||||||
| 截顶蔷薇珊瑚 Montipora truncata | + | 热敏感型 | ||||||
| 膨胀蔷薇珊瑚 Montipora turgescens | + | + | 热敏感型 | |||||
| 滨珊瑚科 Poritidae | 滨珊瑚属 Porites | 扁枝滨珊瑚Porites andrewsi | + | 热耐受型 | ||||
| 疣滨珊瑚Porites annae | + | 热耐受型 | ||||||
| 扁缩滨珊瑚Porites compressa | + | 热耐受型 | ||||||
| 地衣滨珊瑚Porites lichen | + | + | + | + | 热耐受型 | |||
| 澄黄滨珊瑚Porites lutea | + | + | + | 热耐受型 | ||||
| 角孔珊瑚属 Goniopora | 柱角孔珊瑚Goniopora columna | + | + | 热中度适应型 | ||||
| 大角孔珊瑚Goniopora djiboutiensis | + | + | 热中度适应型 | |||||
| 扁平角孔珊瑚 Goniopora planulata | + | + | 热中度适应型 | |||||
| 细角孔珊瑚Goniopora gracilis | + | 热中度适应型 | ||||||
| 诺福克角孔Goniopora norfolkensis | + | + | 热中度适应型 | |||||
| 伯孔珊瑚属 Bernardpora | 斯氏伯孔珊瑚 Bernardpora stutchburyi | + | + | + | + | 热中度适应型 | ||
| 菌珊瑚科 Agariciidae | 牡丹珊瑚属 Pavona | 十字牡丹珊瑚Pavona decussata | + | + | + | 热耐受型 | ||
| 叶形牡丹珊瑚Pavona frondifera | + | + | 热耐受型 | |||||
| 真叶珊瑚科 Euphylliidae | 盔形珊瑚属 Galaxea | 稀杯盔形珊瑚Galaxea astreata | + | 热耐受型 | ||||
| 丛生盔形珊瑚Galaxea fascicularis | + | + | + | + | 热耐受型 | |||
| 木珊瑚科 Dendrophylliidae | 陀螺珊瑚属 Turbinaria | 盾形陀螺珊瑚 Turbinaria peltata | + | + | + | 热耐受型 | ||
| 坚 实 类 群 | 杯形珊瑚科 Pocilloporidae | 杯形珊瑚属 Pocillopora | 鹿角杯形珊瑚 Pocillopora damicornis | + | 热敏感型 | |||
| 疣状杯形珊瑚 Pocillopora verrucosa | + | 热敏感型 | ||||||
| 沙珊瑚科 Psammocoridae | 沙珊瑚属 Psammocora | 深室沙珊瑚 Psammocora profundacella | + | + | 热耐受型 | |||
| 叶状珊瑚科 Lobophylliidae | 棘星珊瑚属 Acanthastrea | 棘星珊瑚Acanthastrea echinata | + | 热耐受型 | ||||
| 刺叶珊瑚属 Echinophyllia | 粗糙刺叶珊瑚 Echinophyllia aspera | + | 热耐受型 | |||||
| 叶状珊瑚属 Lobophyllia | 菌形叶状珊瑚 Lobophyllia agaricia | + | 热中度适应型 | |||||
| 赫氏叶状珊瑚 Lobophyllia hemprichii | + | 热中度适应型 | ||||||
| 裸肋珊瑚科 Merulinidae | 刺孔珊瑚属 Echinopora | 宝石刺孔珊瑚 Echinopora gemmacea | + | 热耐受型 | ||||
| 薄片刺孔珊瑚 Echinopora lamellosa | + | 热耐受型 | ||||||
| 刺星珊瑚属 Cyphastrea | 锯齿刺星珊瑚 Cyphastrea serailia | + | + | + | + | 热耐受型 | ||
| 菊花珊瑚属 Goniastrea | 梳状菊花珊瑚 Goniastrea pectinata | + | 热中度适应型 | |||||
| 网状菊花珊瑚 Goniastrea retiformis | + | + | + | 热中度适应型 | ||||
| 裸肋珊瑚属 Merulina | 阔裸肋珊瑚Merulina ampliata | + | 热中度适应型 | |||||
| 刺柄珊瑚属 Hydnophora | 腐蚀刺柄珊瑚 Hydnophora exesa | + | + | + | 热耐受型 | |||
| 腔星珊瑚属 Coelastrea | 粗糙腔星珊瑚 Coelastrea aspera | + | + | + | 热中度适应型 | |||
| 盘星珊瑚属 Dipsastraea | 黄癣盘星珊瑚 Dipsastraea favus | + | 热中度适应型 | |||||
| 蜥岛盘星珊瑚 Dipsastraea lizardensis | + | 热中度适应型 | ||||||
| 罗图马盘星珊瑚 Dipsastraea rotumana | + | + | + | + | 热中度适应型 | |||
| 标准盘星珊瑚 Dipsastraea speciosa | + | + | + | + | 热中度适应型 | |||
| 美龙氏盘星珊瑚 Dipsastraea veroni | + | + | + | 热中度适应型 | ||||
| 肠珊瑚属 Leptoria | 弗利吉亚肠珊瑚 Leptoria phrygia | + | 热中度适应型 | |||||
| 扁脑珊瑚属 Platygyra | 肉质扁脑珊瑚 Platygyra carnosa | + | + | + | 热中度适应型 | |||
| 交替扁脑珊瑚 Platygyra crosslandi | + | + | + | 热中度适应型 | ||||
| 精巧扁脑珊瑚 Platygyra daedalea | + | + | + | + | 热中度适应型 | |||
| 中华扁脑珊瑚 Platygyra sinensis | + | + | + | + | 热中度适应型 | |||
| 角蜂巢珊瑚 Favites | 秘密角蜂巢珊瑚 Favites abdita | + | + | + | + | 热中度适应型 | ||
| 板叶角蜂巢珊瑚 Favites complanata | + | + | 热中度适应型 | |||||
| 多弯角蜂巢珊瑚 Favites flexuosa | + | + | 热中度适应型 | |||||
| 海孔角蜂巢珊瑚 Favites halicora | + | + | 热中度适应型 | |||||
| 五边角蜂巢珊瑚 Favites pentagona | + | + | + | + | 热中度适应型 | |||
| 同星珊瑚科 Plesiastreidae | 同星珊瑚属 Plesiastrea | 多孔同星珊瑚 Plesiastrea versipora | + | + | 热中度适应型 | |||
| 双星珊瑚科 Diploastraeidae | 双星珊瑚属 Diploastrea | 同双星珊瑚Diploastrea heliopora | + | 热耐受型 | ||||
| 未定类群 | 小星珊瑚属 Leptastrea | 紫小星珊瑚Leptastrea purpurea | + | + | + | + | 热耐受型 | |
| 横小星珊瑚Leptastrea transversa | + | 热耐受型 | ||||||
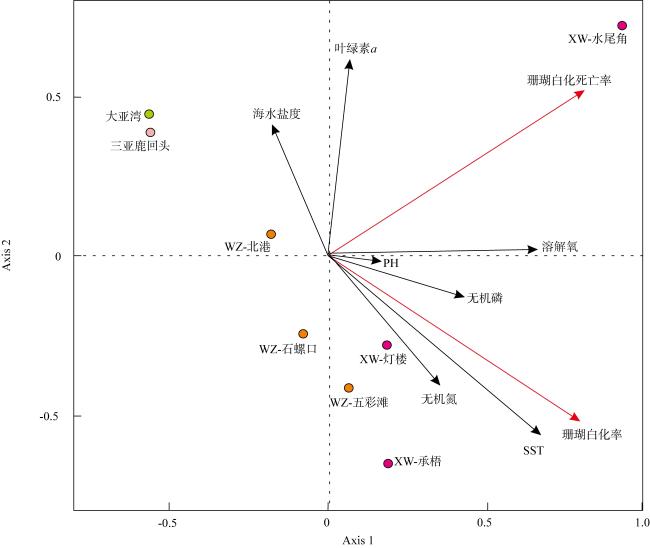
表1 南海北部近岸珊瑚礁区海水环境现因子Tab.1 Sea water environmental parameters in surveyed coral reefs of the northern South China Sea |
| 珊瑚礁区 | SST/℃ | 盐度/‰ | pH | 溶解氧含量/(mg·L-1) | 叶绿素a含量/(μg·L-1) | 无机氮含量/(μmol·L-1) | 无机磷含量/(μmol·L-1) |
|---|---|---|---|---|---|---|---|
| 大亚湾 | 28.9±0.3 | 33.5±0.2 | 8.08±0.12 | 7.71±0.62 | 0.64±0.12 | 1.25±0.31 | 0.05±0.01 |
| 涠洲岛 | 31.3±0.4 | 32.0±0.1 | 8.14±0.09 | 6.97±0.56 | 1.65±0.16 | 4.74±0.64 | 0.18±0.02 |
| 徐闻 | 31.5±0.5 | 33.1±0.1 | 8.14±0.11 | 7.20±0.48 | 5.78±0.45 | 8.96±0.28 | 0.36±0.05 |
| 三亚鹿回头 | 30.1±0.2 | 33.4±0.2 | 8.15±0.14 | 6.87±0.63 | 1.09±0.05 | 9.91±1.52 | 0.05±0.01 |
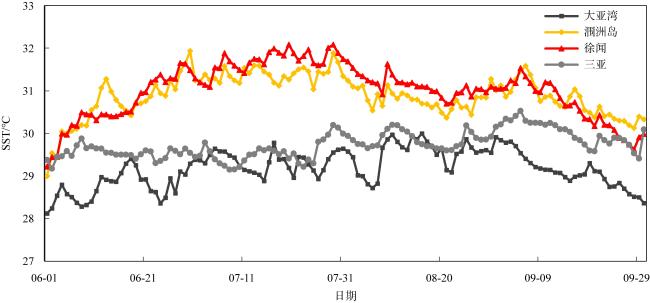
图2 调查区域2020年夏季SST日变化情况2020数据来自欧洲中期天气预报中心全球第五代大气再分析数据集(ERA5), 下载自哥白尼气候变化服务(Copernicus Climate Change Service, C3S)气候数据存储(Climate Data Store, CDS), 链接: https://cds.climate.copernicus.eu/cdsapp#!/dataset/reanalysis-era5-single-levels?tab=form Fig. 2 Sea surface temperatures (SSTs) in the study region from summer |
表2 南海北部近岸不同珊瑚礁区的珊瑚优势种组成及其白化率Tab. 2 Bleaching status of dominant scleractinian coral species on coral reefs of the northern South China Sea |
| 珊瑚礁区 | 造礁石珊瑚优势种 | 重要值百分比/% | 覆盖度/% | 白化率/% | 耐热性类型 | |
|---|---|---|---|---|---|---|
| 大亚湾珊瑚栖息地 | 澄黄滨珊瑚 | Porites lutea | 15.4 | 3.55 | 0 | 热耐受型 |
| 锯齿刺星珊瑚 | Cyphastrea serailia | 11.0 | 1.94 | 0 | 热耐受型 | |
| 翼形蔷薇珊瑚 | Montipora peltiformis | 9.6 | 3.15 | 0 | 热敏感型 | |
| 疣滨珊瑚 | Porites annae | 6.6 | 1.74 | 0 | 热耐受型 | |
| 单独鹿角珊瑚 | Acropora solitaryensis | 6.2 | 1.95 | 0 | 热敏感型 | |
| 罗图马盘星珊瑚 | Dipsastraea rotumana | 6.0 | 0.86 | 0 | 热中度适应型 | |
| 涠洲岛珊瑚礁 | 十字牡丹珊瑚 | Pavona decussata | 15.9 | 3.93 | 19.08 | 热耐受型 |
| 澄黄滨珊瑚 | Porites lutea | 15.8 | 4.22 | 74.64 | 热耐受型 | |
| 薄片刺孔珊瑚 | Echinopora lamellosa | 13.0 | 4.39 | 21.64 | 热耐受型 | |
| 秘密角蜂巢珊瑚 | Favites abdita | 5.8 | 0.59 | 89.83 | 热中度适应型 | |
| 徐闻珊瑚礁 | 柱角孔珊瑚 | Goniopora columna | 13.7 | 2.94 | 95.24 | 热中度适应型 |
| 斯氏伯孔珊瑚 | Bernardpora stutchburyi | 13.5 | 1.96 | 98.47 | 热中度适应型 | |
| 盾形陀螺珊瑚 | Turbinaria peltata | 10.2 | 1.65 | 84.85 | 热耐受型 | |
| 澄黄滨珊瑚 | Porites lutea | 8.4 | 1.15 | 93.04 | 热耐受型 | |
| 深室沙珊瑚 | Psammocora profundacella | 6.2 | 1.66 | 91.57 | 热耐受型 | |
| 三亚鹿回头珊瑚礁 | 澄黄滨珊瑚 | Porites lutea | 16.6 | 8.88 | 0 | 热耐受型 |
| 丛生盔形珊瑚 | Galaxea fascicularis | 7.8 | 3.23 | 0 | 热耐受型 | |
| 斯氏伯孔珊瑚 | Bernardpora stutchburyi | 7.3 | 3.23 | 0 | 热中度适应型 | |
| 繁锦蔷薇珊瑚 | Montipora efflorescens | 7.2 | 4.11 | 0 | 热敏感型 | |
| 膨胀蔷薇珊瑚 | Montipora turgescens | 5.5 | 2.37 | 0 | 热敏感型 | |
| 鹿角杯形珊瑚 | Pocillopora damicornis | 5.2 | 1.55 | 0 | 热敏感型 | |
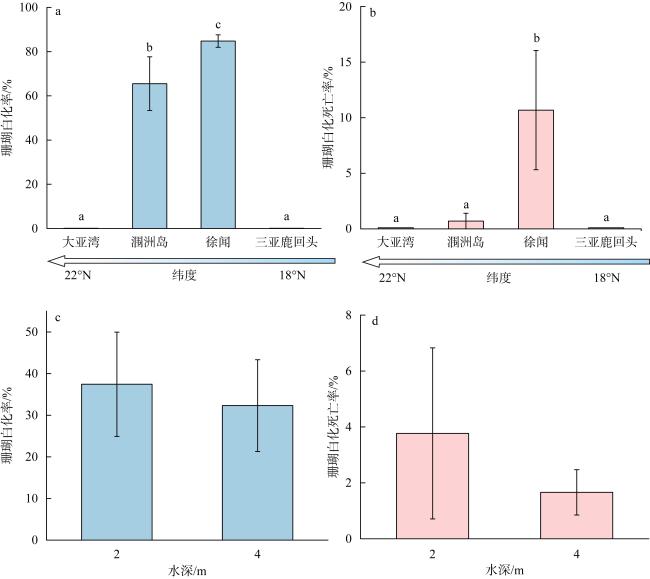
图4 珊瑚白化率与珊瑚白化死亡率在各珊瑚礁区和水深之间的变化柱状图内不同字母标记表明存在显著性差异(P<0.05) Fig. 4 Changes of coral bleaching and bleaching mortality percentages among coral reefs and water depths |
图6 涠洲岛和徐闻珊瑚礁区珊瑚的热白化照片a. 霜鹿角珊瑚; b. 薄片刺孔珊瑚; c. 十字牡丹珊瑚; d. 盾形陀螺珊瑚; e. 粗糙刺叶珊瑚; f. 柱角孔珊瑚(左)和斯氏伯孔珊瑚(右); g. 澄黄滨珊瑚; h. 肉质扁脑珊瑚; i. 秘密角蜂巢珊瑚。其中a为枝状珊瑚; b、c为叶状珊瑚; d为板状珊瑚; e、f为皮壳状珊瑚; g—i为块状珊瑚 Fig. 6 Bleaching scleractinian corals on Weizhou Island and Xuwen coral reefs. (a) Acropora pruinose; (b) Echinopora lamellose; (c) Pavona decussate; (d) Turbinaria peltate; (e) Echinophyllia aspera; (f) Goniopora columna (left) and Bernardpora stutchburyi (right); (g) Porites lutea; (h) Platygyra carnosus; (i) Favites abdita |
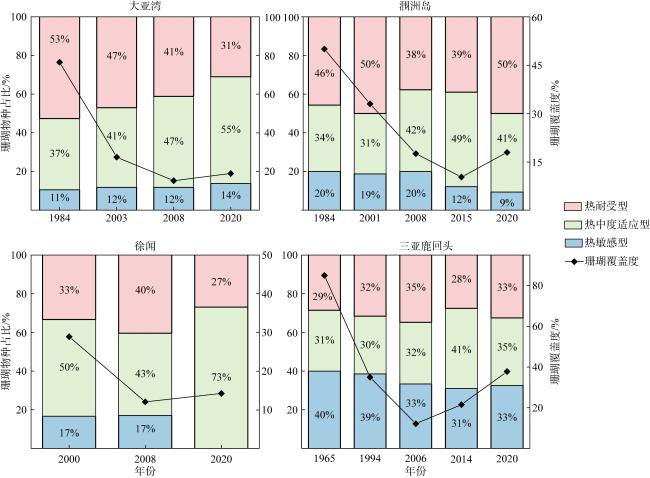
图7 南海北部造礁石珊瑚群落的耐热性组成历史演变数据来源于本研究和历史文献, 包括张元林 等(1987); 于登攀 等(1996); 赵焕庭 等(2002); 卢伙胜 等(2003); 林昭进 等(2007); 赵美霞 等(2009); Chen等(2009); 赵美霞 等(2010); 黄晖 等(2011); 孙有方 等(2018); Yu等(2019); 周浩郎 等(2020) Fig. 7 Evolution of heat tolerance composition for coral community structure in the northern South China Sea |
| [1] |
郭峰, 肖家光, 田鹏, 等, 2022. 大亚湾及大鹏半岛沿岸造礁石珊瑚现状与生态脆弱性评价[J]. 应用海洋学学报, 41(4): 568-582.
|
| [2] |
国家海洋局, 2005. 珊瑚礁生态监测技术规程, HY/T 082-2005[S]. 北京: 中国标准出版社: 1-20.
State Oceanic Administration, 2005. Technical specification for eco-monitoring of coral reef ecosystem, HY/T 082-2005[S]. Beijing: Standards Press of China: 1-20 (in Chinese).
|
| [3] |
黄晖, 张浴阳, 练健生, 等, 2011. 徐闻西岸造礁石珊瑚的组成及空间分布[J]. 生物多样性, 19(5): 505-510.
|
| [4] |
黄猛, 2007. 广东省海洋与渔业自然保护区建设概况[J]. 海洋与渔业, (10): 14-16 (in Chinese).
|
| [5] |
江绿苗, 陈天然, 赵宽, 等, 2023. 南海北部涠洲岛边缘珊瑚礁的生物侵蚀实验研究[J/OL]. 热带海洋学报. [2023-04-17]. https://doi.org/10.11978/2023002.
|
| [6] |
李禹辉, 邱云, 杨龙奇, 等, 2021. 大亚湾及其邻近海域夏季温度、盐度的分布特征[J]. 应用海洋学学报, 40(2): 284-292.
|
| [7] |
梁文, 周浩郎, 王欣, 等, 2021. 涠洲岛西南部海域造礁石珊瑚的群落结构特征分析[J]. 海洋学报, 43(11): 123-135.
|
| [8] |
林昭进, 邱永松, 张汉华, 等, 2007. 大亚湾浅水石珊瑚的分布现状及生态特点[J]. 热带海洋学报, 26(3): 63-67.
|
| [9] |
卢伙胜, 何秀玲, 陈春亮, 等, 2003. 广东徐闻西部沿岸海区“珊瑚类”的物种及其分布[J]. 台湾海峡, 22(4): 445-448.
|
| [10] |
施祺, 赵美霞, 黄玲英, 等, 2010. 三亚鹿回头岸礁区人类活动及其对珊瑚礁的影响[J]. 热带地理, 30(5): 486-490, 509.
|
| [11] |
孙有方, 雷新明, 练健生, 等, 2018. 三亚珊瑚礁保护区珊瑚礁生态系统现状及其健康状况评价[J]. 生物多样性, 26(3): 258-265.
|
| [12] |
唐灵, 聂宇华, 王平, 等, 2022. 1974—2020年珠江口外海海洋热浪变化趋势分析[J]. 热带海洋学报, 41(6): 143-150.
|
| [13] |
吴莹莹, 雷新明, 黄晖, 等, 2021. 南海典型珊瑚礁生态系统健康评价方法研究[J]. 热带海洋学报, 40(4): 84-97.
|
| [14] |
杨振雄, 张敬怀, 吕向立, 等, 2021. 涠洲岛造礁石珊瑚群落变化特征及其环境影响因子[J]. 生态学报, 41(18): 7168-7179.
|
| [15] |
于登攀, 邹仁林, 1996. 鹿回头造礁石珊瑚群落多样性的现状及动态[J]. 生态学报, 16(6): 559-564.
|
| [16] |
张文静, 郑兆勇, 张婷, 等, 2020. 1960-2017年北部湾珊瑚礁区海洋热浪增强原因分析[J]. 海洋学报, 42(5): 41-48.
|
| [17] |
张元林, 邹仁林, 1987. 大亚湾浅水石珊瑚群落结构的研究[J]. 热带海洋, 6(1): 12-18.
|
| [18] |
赵焕庭, 王丽荣, 宋朝景, 等, 2002. 雷州半岛灯楼角珊瑚岸礁的特征[J]. 海洋地质与第四纪地质, 22(2): 35-40.
|
| [19] |
赵美霞, 余克服, 张乔民, 等, 2009. 近50a来三亚鹿回头石珊瑚物种多样性的演变特征及其环境意义[J]. 海洋环境科学, 28(2): 125-130.
|
| [20] |
赵美霞, 余克服, 张乔民, 等, 2010. 近50年来三亚鹿回头岸礁活珊瑚覆盖率的动态变化[J]. 海洋与湖沼, 41(3): 440-447.
|
| [21] |
中国国家标准化管理委员会, 2007. 海洋调查规范第4部分:海水化学要素调查, GB/T 12763. 4-2007[S]. 北京: 中国标准出版社: 1-65.
Standardization Administration of the People's Republic of China, 2007, Specifications for oceanographic survey part: survey of chemical parameters in sea water, GB/T 12763. 4-2007 [S]. Beijing: Standards Press of China: 1-65 (in Chinese).
|
| [22] |
周浩郎, 王欣, 梁文, 2020. 涠洲岛珊瑚礁特点、演变及保护与修复对策的思考[J]. 广西科学院学报, 36(3): 228-236.
|
| [23] |
邹仁林, 2001. 中国动物志:腔肠动物门, 珊瑚虫纲, 石珊瑚目, 造礁石珊瑚[M]. 北京: 科学出版社: 1-289 (in Chinese).
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}