
Journal of Tropical Oceanography >
Response of dissimilatory nitrate reduction processes in coral reef sediments of the Weizhou island to temperature changes
Copy editor: SUN Cuici
Received date: 2023-08-02
Revised date: 2023-09-13
Online published: 2023-11-09
Supported by
Guangxi Natural Science Foundation(2019GXNSFAA185001)
College Students’ Innovative Entrepreneurial Training Plan Program(S202210593227)
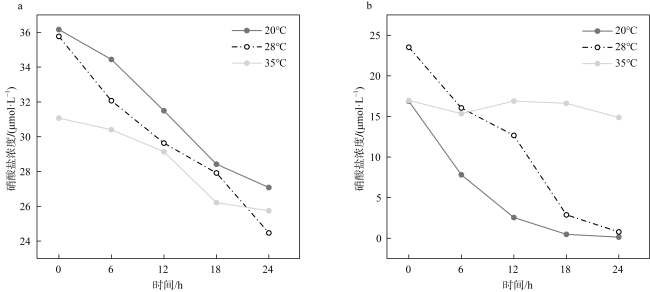
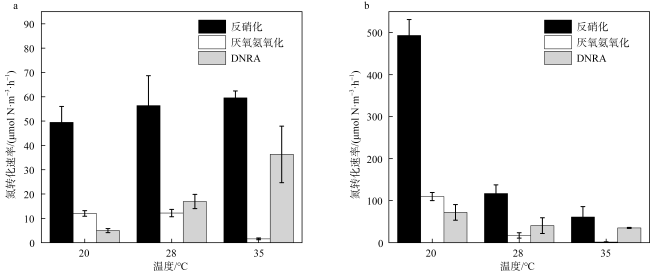
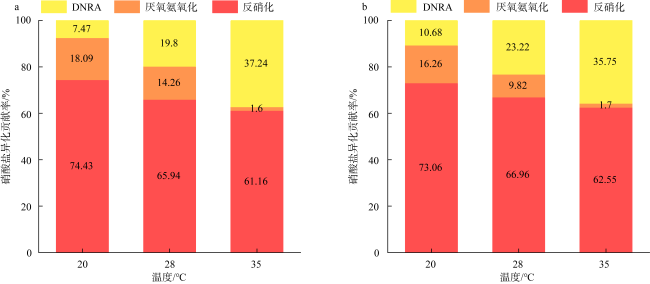
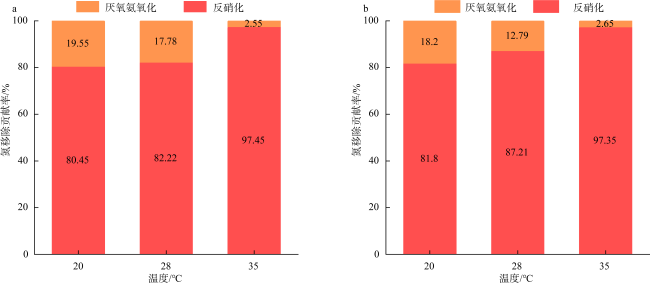
Dissimilatory nitrate reduction processes, including denitrification, anaerobic ammonium oxidation, and dissimilatory nitrate reduction to ammonium (DNRA) regulate the removal and retention of nitrogen elements in the ocean. Global warming affecting the nitrogen cycling processes in sediments, but it is not yet clear the differences in response of dissimilatory nitrate reduction processes in carbonate sand and silicate sand sediments to increased temperature. Therefore, we used 15N isotope tracing technology and temperature-controlled incubation of sediments to investigate the response of dissimilatory nitrate reduction processes in different sediment types in the coral reef area of the Weizhou Island to increased temperature. The results indicate that DNRA and denitrification processes exhibited significant differences in response to increased temperature in two different types of sediments. The rates of denitrification and DNRA in silicate sand were significantly increased with the increased temperature, but those in carbonate sand exhibited opposite trend, which may be related to differences in microbial community in different sediments. Due to anaerobic ammonium oxidation process being more sensitive to increased temperature, the contribution of denitrification to nitrogen removal increased, but its byproduct N2O may have a negative impact on climate. In addition, the increase in temperature led to an increase in the contribution of DNRA to nitrate dissimilation reduction, which will exacerbate the negative impact of nitrogen excess on the coral reef ecosystem of the Weizhou Island.
Key words: denitrification; increased temperature; DNRA; sediments; coral reef
MO Danyang , NING Zhiming , YANG Bin , XIA Ronglin , LIU Zhijin . Response of dissimilatory nitrate reduction processes in coral reef sediments of the Weizhou island to temperature changes[J]. Journal of Tropical Oceanography, 2024 , 43(4) : 137 -143 . DOI: 10.11978/2023108
| [1] |
刘祯, 郑志宏, 2022. 水体中DNRA与反硝化之间竞争研究进展[J]. 河南化工, 39(9): 4-7.
|
| [2] |
罗跃辉, 阮晓红, 李荣富, 等, 2017. 太湖西部湖区沉积物厌氧氨氧化潜在速率及其脱氮贡献研究[J]. 环境科学学报, 37(11): 4187-4194.
|
| [3] |
亚涛, 2022. 温度对厌氧氨氧化系统运行特性及微生物相互作用关系的影响机理研究[D]. 北京: 北京化工大学.
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}