
Journal of Tropical Oceanography >
Microbial communities and specific strains within beachrocks of the South China Sea: implications for the origin of beachrock*
Copy editor: SUN Cuici
Received date: 2023-10-07
Revised date: 2023-10-23
Online published: 2023-12-08
Supported by
National Key Research and Development Program of China(2021YFC3100603)
National Key Research and Development Program of China(2022YFC3102402)
National Natural Science Foundation of China(42306076)
National Natural Science Foundation of China(42376165)
Key Research and Development Program of Hainan Province(ZDYF2022SHFZ072)
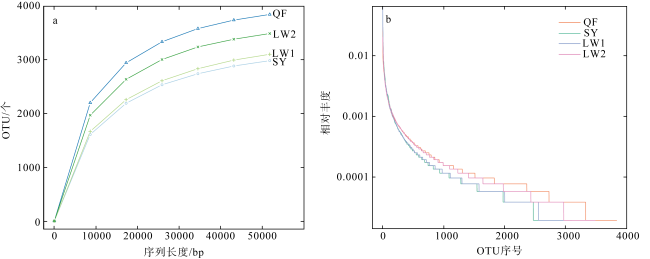
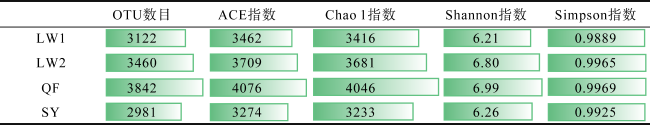
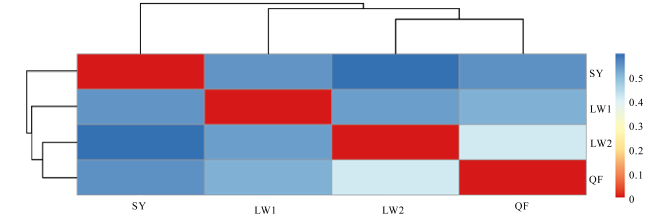
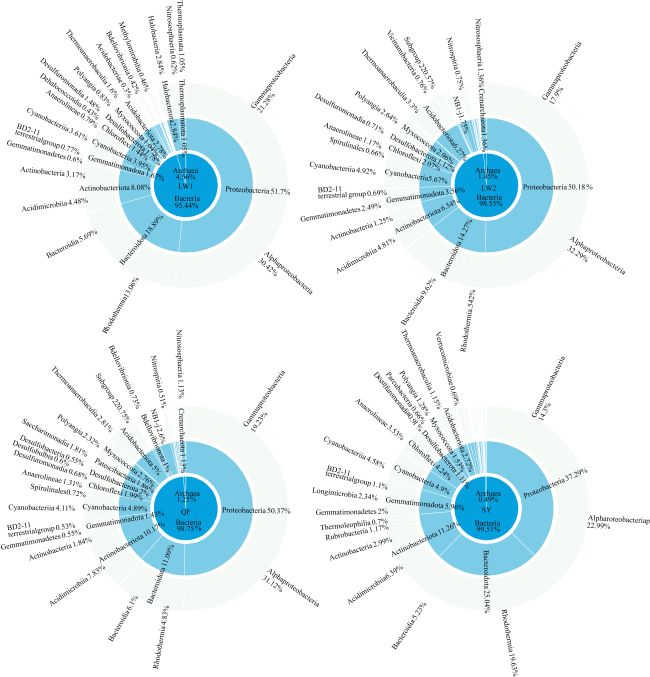
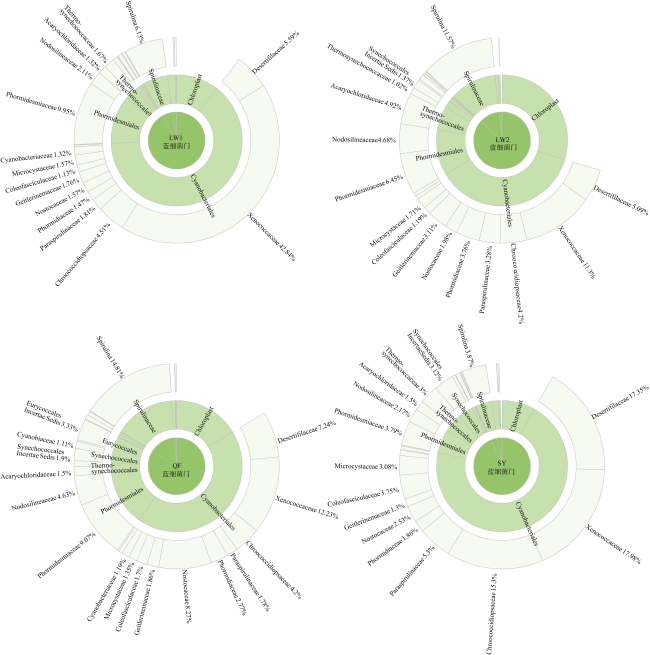
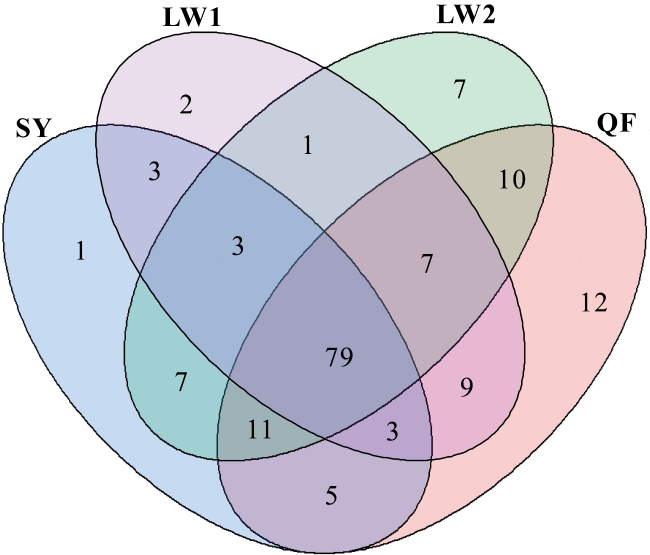
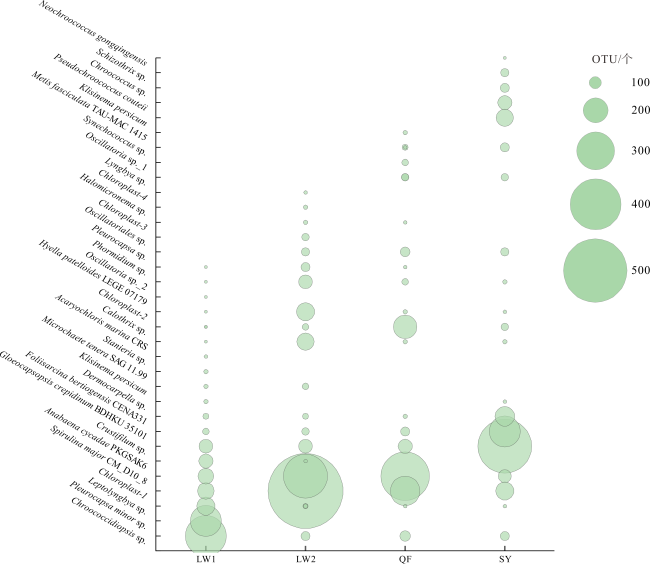
Abundant bacterial-algal microorganisms and their metabolisms play important roles in early cementation of beachrocks along intertidal-supratidal zones of tropical-subtropical shorelands. In this study, 16S rRNA high-throughput sequencing analysis was employed to reveal the compositions of prokaryotic microbial communities and identify the common strains/microbial functional groups that dominated the beachrocks’ cementation. The Quanfu turf algae-bearing beachrocks have the highest biodiversity. The diversity of Longwan beachrocks in upper intertidal zone is higher than that in lower intertidal zone. The prokaryotic biodiversity of consolidated Shiyu beachrocks is the lowest. The bacterial phylum in the four sampling localities was roughly similar, which includes Proteobacteria, Bacteroideta, Actinobacteria, Cyanobacteria, Chloroflexi, Gemmatimonadetes and Desulfobacterota in descending order. The main cyanobacteria species were same. The common calcified cyanobacterial strain-Calothrix sp. was all detected, and the extracellular polymer substances could be the hotspots for precipitation of carbonate biomaterials. Denitrobacteria and sulfate-reducing bacteria detected in microbial communities indicate the anaerobic environment and steep redox gradient below the superficial beachrocks. Strong photosynthesis, sulfate reduction and denitrification in particle surface and intergranular pore together significantly rise pH, and promote the nucleation and growth of carbonate biomaterials, accelerating the consolidation of loose beach sands.
Key words: prokaryote; community structure; cyanobacteria; beachrock; South China Sea
LIU Didi , ZHANG Xiyang , SUN Fulin , WANG Mingzhuang , TAN Fei , SHI Qi , WANG Guan , YANG Hongqiang . Microbial communities and specific strains within beachrocks of the South China Sea: implications for the origin of beachrock*[J]. Journal of Tropical Oceanography, 2024 , 43(4) : 112 -122 . DOI: 10.11978/2023142
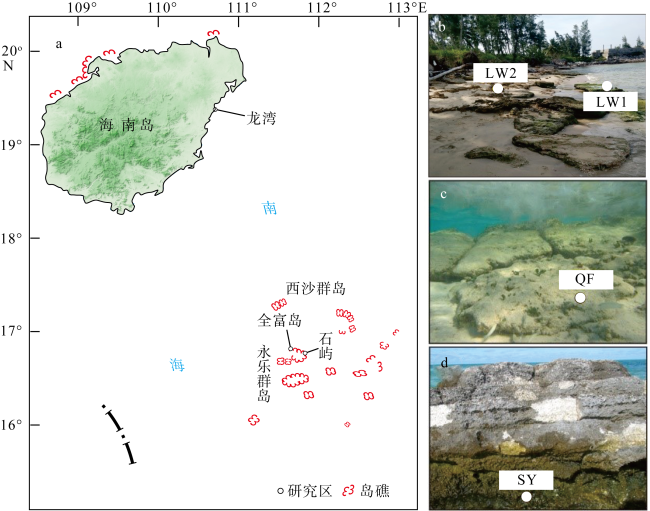
图1 a. 采样点区域位置图; b. 为龙湾海滩岩野外照片; c. 全富岛海滩岩样品(QF)采自海滩岩表面草皮海藻; d. 石屿海滩岩样品(SY)为潮间带海滩岩表面该图基于海南测绘地理信息局的审图号为琼S(2021)120号的标准地图制作, 底图无修改。龙湾1号取样点海滩岩样品(LW1)位于潮间带下部, 龙湾2号取样点海滩岩样品(LW2)位于潮间带上部 Fig. 1 (a) Map of sampling sites; (b) LW1 beachrocks from the lower part of intertidal zone and LW2 beachrocks from the upper part of intertidal zone; (c) QF beachrock and the surficial turf algae; (d) SY beachrock from intertidal zone |
| [1] |
白洁, 李海艳, 赵阳国, 2009. 黄海北部不同站位海洋细菌群落分布特征[J]. 微生物学报, 49(3): 343-350.
|
| [2] |
白洁, 刘小沙, 侯瑞, 等, 2014. 南海南部海域浮游细菌群落特征及影响因素研究[J]. 中国环境科学, 34(11): 2950-2957.
|
| [3] |
陈倩茹, 夏雪, 王川, 等, 2021. 沙颍河下游城市黑臭内河沉积物微生物群落季节变化特征[J]. 水生生物学报, 45(1): 182-189.
|
| [4] |
高旭波, 潘振东, 龚培俐, 等, 2022. 微生物诱导碳酸盐岩沉淀过程及作用机理[J]. 中国岩溶, 41(3): 441-452.
|
| [5] |
李建生, 冯炎基, 1998. 华南海滩岩及若干问题讨论[J]. 矿物岩石地球化学通报, 17(3): 45-47.
|
| [6] |
李平日, 1988. 华南全新世海滩岩及其古地理意义[J]. 海洋地质与第四纪地质, 8(4): 21-29.
|
| [7] |
仇潇洒, 2020. 陶厄氏菌属(Thauera)富集的SBR系统脱氮性能研究[D]. 西安: 长安大学: 1-65.
|
| [8] |
孙金龙, 徐辉龙, 2009. 中国的海滩岩研究与进展[J]. 热带海洋学报, 28(2): 103-108.
|
| [9] |
滕建彬, 金春花, 沈建伟, 2007a. 海南岛鹿回头水尾岭海滩岩中的微生物岩[J]. 海洋地质与第四纪地质, 27(3): 25-30.
|
| [10] |
滕建彬, 沈建伟, 2007b. 海南岛鹿回头水尾岭海滩岩中的微生物碳酸盐沉积研究[J]. 中国科学D辑: 地球科学, 37(10): 1338-1348.
|
| [11] |
王月, 沈建伟, 龙江平, 2011. 海南岛三亚小东海珊瑚礁坪生态沉积分带和碳酸盐沉积作用[J]. 中国科学D辑: 地球科学, 41(3): 362-374.
|
| [12] |
朱长歧, 周斌, 刘海峰, 2015. 南海海滩岩的细观结构及其基本物理力学性质研究[J]. 岩石力学与工程学报, 34(4): 683-693.
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}