
Journal of Tropical Oceanography >
Diversity of culturable bacteria in the sedimentary sands of the Meiji Reef
Editor: LIN Qiang
Received date: 2023-12-05
Revised date: 2024-02-21
Online published: 2024-02-26
Supported by
Hainan Provincial Natural Science Foundation of China(321CXTD447)
National Natural Science Foundation of China(32170018)
Science and Technology Planning Project of Guangdong Province of China(2021B1212050023)
The special environment of reefs in the South China Sea has created a rich and colorful microbial world, where a large number of microorganisms are yet to be excavated and utilized. Due to geographical conditions and other factors, there are few studies on them. In this paper, culturable microorganisms were selectively isolated from the sedimentary sands of the Meiji Reef by using oligotrophic media combined with high salt and high temperature conditions. And their diversity and distribution were also studied, a variety of functional media were used to explore the functions of these microorganisms. The 674 isolates belong to 5 phyla, 6 classes, 26 orders, 38 families, 62 genera, 146 species, respectively, and 42 potential new species with less than 98.65% similarity composes around 28.7% all of them. Among the cultured bacteria obtained, the Phyla of Actinobacteriota and Pseudomonadota are the most abundant, accounting for 61.6% of the total isolated strains. Functional analysis revealed that 2 species have the phospho-reducing activity, 36 species have the urease-producing activity and 57 species have the iron-bearing activity. The research results indicate that there are microbial abundance, much new species resources and high diversity of microorganism with special ecology functions in the reef environment. In study, these accumulates rich microbial species resources, laying the foundation for the deeply exploration of microbial resources utilization and the ecological restoration of the coral-sands ecosystem in the Meiji Reef.
JIANG Rouyun , JIAN Lili , SHI Songbiao , TIAN Xinpeng . Diversity of culturable bacteria in the sedimentary sands of the Meiji Reef[J]. Journal of Tropical Oceanography, 2024 , 43(6) : 170 -180 . DOI: 10.11978/2023183
表1 培养基类型及其组成成分Tab. 1 Typic media and their components used in this study |
| 培养基 | 成分 |
|---|---|
| R2A | 胰蛋白胨0.25g, 可溶性淀粉0.5g, 酵母浸粉0.5g, K2HPO4 0.3g, MgSO4 0.1g, 丙酮酸钠0.3g, 蛋白胨0.25g, 葡萄糖0.5g, 酸水解酪蛋白0.5g, 稀释20倍, 琼脂15g, 水1L |
| 2216E | 酵母浸粉1.0g, 柠檬酸铁0.1g, NaCl 19.45g, MgCl2 5.98g, Na2SO4 3.24g, CaCl2 1.8g, KCl 0.55g, Na2CO3 0.16g, KBr 0.08g, SrCl2 0.034g, H3BO3 0.022g, Na2O·SiO2 0.004g, NaF 0.002 4g, NH4NO3 0.001 6g, Na2HPO4 0.008g, 蛋白胨5.0g, 稀释20倍, 琼脂15g, 水1L |
| SN | NaNO3 0.75g, K2HPO4 0.015 9g, EDTA二钠0.005 6g, Na2CO3 0.010 4g, VitaminB12 0.001g(过滤除菌), 1×10-6微量盐(乙酸6.25g, 柠檬酸铁铵6.0g, Na2MoO4·2H2O 0.39g, MnCl2·4H2O 1.4g, Co(NO3)2·6H2O 0.025g, ZnSO3·7H2O 0.222g, 单独灭菌), 琼脂15g, 海水500mL, 水500mL |
| R | NaCl 25g, MgSO4·7H2O 9g, CaCl2·2H2O 0.14g, KCl 0.7g, Na2HPO4·12H2O 0.25g, Na2-EDTA 30mg, H3BO3 34mg, FeSO4·7H2O 10mg, FeCl3·6H2O 1.452mg, MnCl2·4H2O 4.32mg, CoCl2·6H2O 0.12mg, Na2MoO·2H2O 0.63mg, RbCl 0.141 5mg, LiCl 0.61mg, NaBr 6.4mg KI 0.006 55mg, SrCl2·6H2O 3.04mg, V2O5 0.001 785mg, ZnCl2 0.312mg, RPMI 500mg, EagleMedium 500mg, L-glutamine 15mg, NaHCO3 100mg, 海盐18g, 琼脂15g, 水1L |
| 无机磷固体 | MnSO4·4H2O 0.03g, (NH4)2SO4 0.5g, NaCl 0.3g, MgSO4·7H2O 0.3g, KCl 0.3g, FeSO4·4H2O 0.036g, 葡萄糖10g, 有机磷植酸钙2.0g [或无机磷Ca3(PO4)210g], 水1L, 琼脂15g, pH 7.0 |
| 亚历山大硅酸盐 | 蔗糖5g, Na2HPO4 2g, FeCl3 0.005g, CaSO4 0.1g, MgSO4·7H2O 0.5g琼脂15g, 水1L, pH 7.0~7.5 |
| DF | KH2PO4 4g, MgSO4·7H2O 0.2g, 葡萄糖2g, 葡萄糖酸2g, 柠檬酸2g, Na2HPO4 6g, H3BO3 0.01mg, MgSO4 0.011 2mg, ZnSO4 0.124 6 mg, CuSO4 0.078 2mg, MoO3 0.01mg, 水1L, 琼脂15g, pH 7.0 |
| ADF | KH2PO4 4g, Na2HPO4 6g, (NH4)2SO4 2g, MgSO4·7H2O 0.2g, 葡萄糖2g, 葡萄糖酸2g, 柠檬酸2g, H3BO3 0.01mg, MgSO4 0.011 2 mg, ZnSO4 0.124 6 mg, MoO3 0.01mg, 1-氨基羰酰-1-环丙烷羧酸(ACC)5mmol/L, FeSO4·7H2O 0.1mg, CuSO4 0.078 2mg, 水1L, 琼脂15g, pH 7.0 |
| CAS | CAS 0.605g, Na2HPO4·12H2O 1.213 5g, HDTMA 0.0729g, NaH2PO4·2H2O 0.295 25g, FeCl3·6H2O 0.000 3g, NHCl4 0.125g, KH2PO4 0.037 5g, NaCl 0.625g, 琼脂9g, 水1L, pH 6.7~6.9 |
| 产脲酶 | 蛋白胨1g, NaCl l5g, KH2PO4 2g, 葡萄糖1g, 酚红0.012g, 琼脂15g, 水1L, pH 7.0 |
| 三丁酸甘油酯 | 三丁酸甘油酯40mL,阿拉伯树胶4g, 琼脂15g, 水1L, pH 7.0 |
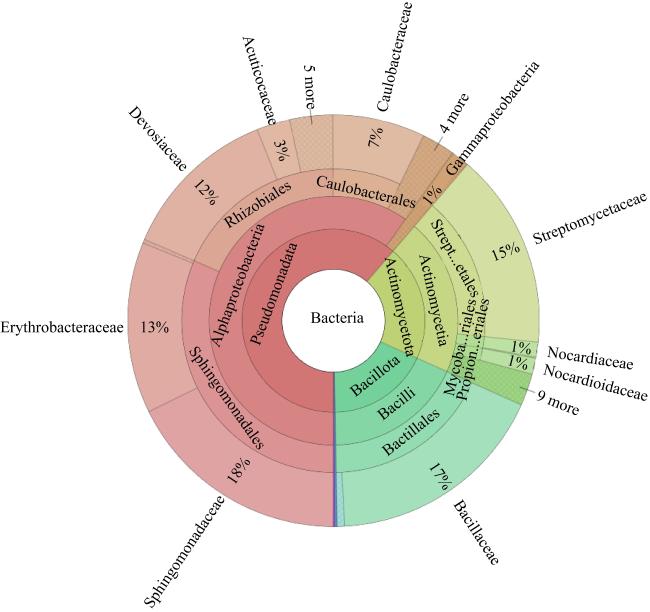
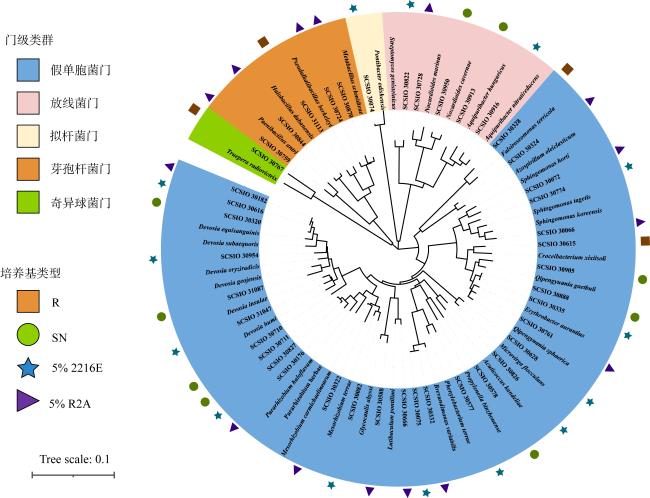
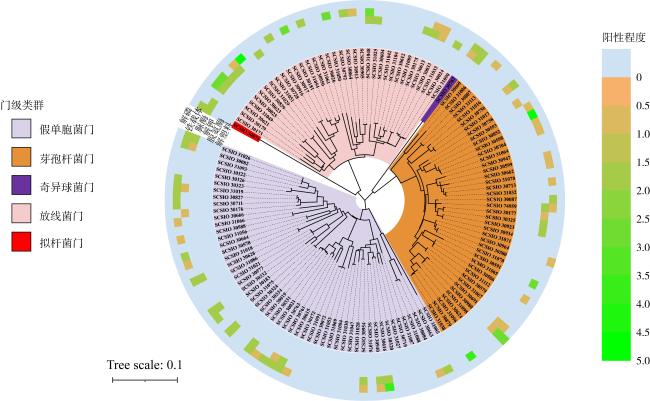
图1 纯培养菌株在不同分类等级的分布情况Bacteria: 细菌域; Actinomycetota: 放线菌门; Pseudomonadata: 变形菌菌门; Bacillota: 芽孢杆菌门; Alphaproteobacteria: α-变形菌纲; Gammaproteobacteria: γ-变形菌纲; Actinomycetia: 放线菌纲; Bacilli: 芽孢杆菌纲; Rhizobiales: 根瘤菌目; Mycobacteriales: 核菌目; Streptomycetales: 链霉菌目; Propionibacteriales: 丙酸菌目; Sphingomonadales: 鞘氨醇单胞菌目; Streptosporangiaceae: 链孢菌科; Nocardiaceae: 诺卡菌科; Devosiaceae: 德沃斯菌科; Erythrobacteraceae: 动孢红杆菌科; Nocardioidaceae: 类诺卡菌科; 链霉菌科Streptomycetaceae; Sphingomonadaceae: 鞘氨醇单胞菌科; Acuticoccaceae: 锐球菌科 Fig. 1 The distribution of pure culture strains in different taxonomic clades |
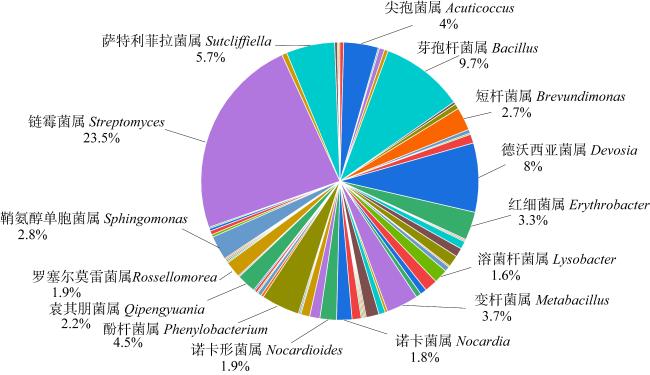
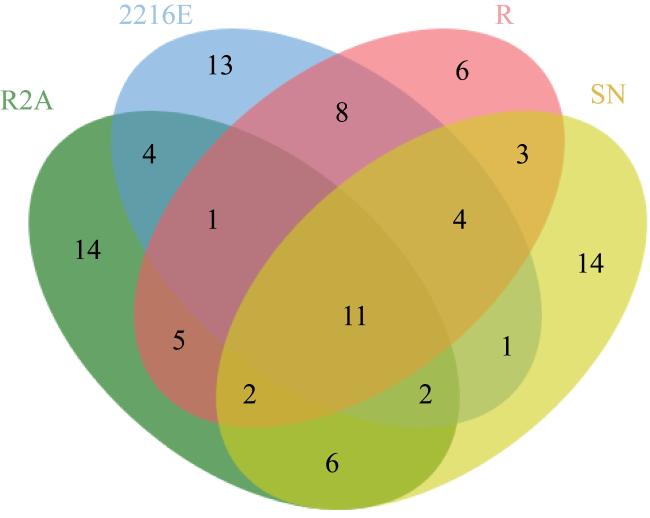
图3 寡营养条件下5个样品获得的可培养物种多样性Fig. 3 Diversity of culturable species obtained from 5 samples under the oligotrophic condition |
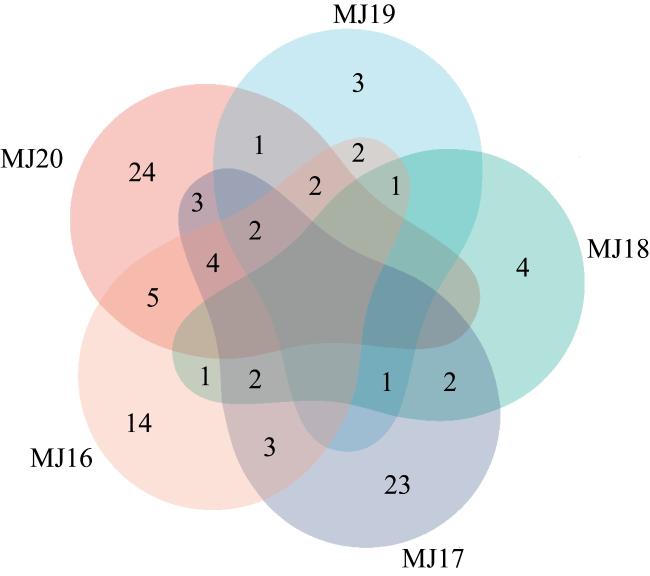
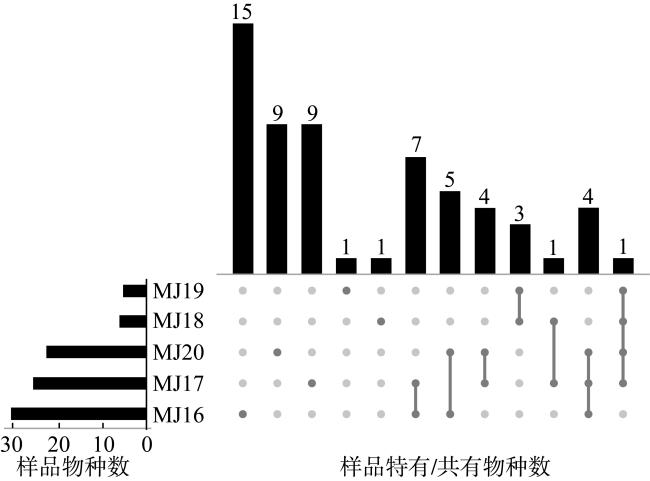
表2 不同条件下5个样品获得的微生物多样性情况Tab. 2 Microbial diversity obtained from 5 samples under different conditions |
| 条件 | 样品 | 门 | 纲 | 目 | 科 | 属 | 种 | 株 |
|---|---|---|---|---|---|---|---|---|
| 总条件 | MJ16 | 3 | 4 | 14 | 20 | 30 | 54 | 178 |
| MJ17 | 5 | 6 | 15 | 24 | 35 | 57 | 157 | |
| MJ18 | 3 | 4 | 6 | 7 | 9 | 14 | 56 | |
| MJ19 | 3 | 4 | 5 | 7 | 12 | 18 | 50 | |
| MJ20 | 4 | 5 | 16 | 19 | 29 | 53 | 233 | |
| 寡营养 | MJ16 | 3 | 4 | 11 | 16 | 26 | 36 | 103 |
| MJ17 | 5 | 6 | 14 | 20 | 27 | 40 | 100 | |
| MJ18 | 3 | 4 | 6 | 7 | 8 | 11 | 33 | |
| MJ19 | 3 | 3 | 4 | 6 | 10 | 12 | 27 | |
| MJ20 | 4 | 5 | 14 | 17 | 26 | 41 | 159 | |
| 高盐(150‰ NaCl) | MJ16 | 3 | 3 | 10 | 14 | 20 | 31 | 73 |
| MJ17 | 3 | 3 | 8 | 11 | 15 | 26 | 51 | |
| MJ18 | 3 | 4 | 4 | 4 | 5 | 6 | 8 | |
| MJ19 | 3 | 3 | 3 | 3 | 4 | 6 | 7 | |
| MJ20 | 3 | 4 | 12 | 13 | 17 | 24 | 60 | |
| 高温(50℃) | MJ16 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| MJ17 | 1 | 1 | 1 | 1 | 1 | 2 | 6 | |
| MJ18 | 1 | 1 | 1 | 1 | 1 | 3 | 15 | |
| MJ19 | 1 | 1 | 1 | 1 | 1 | 3 | 16 | |
| MJ20 | 2 | 2 | 2 | 2 | 3 | 3 | 14 |
表3 高温条件下5个样品获得的物种情况 |
Tab. 3 Strains obtained from 5 samples under the high temperature condition |
| 样品 | 纲 | 种或亚种 |
|---|---|---|
| MJ16 | 芽孢杆菌纲Bacilli | Cytobacillus pseudoceanisediminis |
| 放线菌纲Actinomycetes | Streptomyces radiopugnans | |
| MJ17 | 芽孢杆菌纲Bacilli | Bacillus altitudinis、Bacillus tequilensis |
| MJ18 | 芽孢杆菌纲Bacilli | Bacillus altitudinis、Bacillus xiamenensis、Bacillus tequilensis |
| MJ19 | 芽孢杆菌纲Bacilli | Bacillus altitudinis、Bacillus xiamenensis、Bacillus tequilensis、Bacillus safensis subsp. safensis |
| MJ20 | 芽孢杆菌纲Bacilli | Fredinandcohnia onubensis、Paenibacillus cisolokensis |
| 放线菌纲Actinomycetes | Streptomyces radiopugnans |
图5 寡营养条件下4种培养基获得可培养物种的数量和分布Fig. 5 Distribution of culturable species obtained by four selective media under oligotrophic condition |
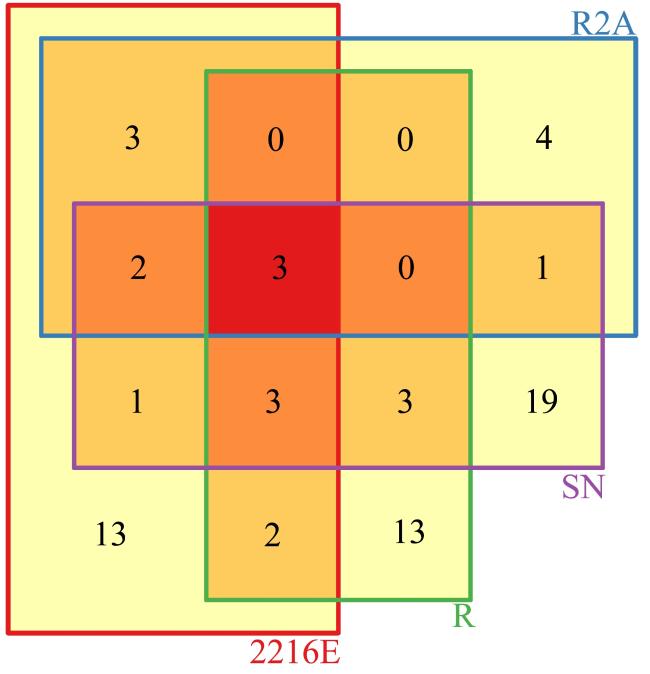
图6 高盐(150‰ NaCl)条件下4种培养基获得的物种分布情况Fig. 6 Distribution of species obtained from four selective media under 150‰ NaCl high salt condition |
表4 不同分离培养基获得的微生物多样性情况Tab. 4 Microbial diversity obtained from the isolation media under different conditions |
| 条件 | 样品 | 门 | 纲 | 目 | 科 | 属 | 种 | 株 |
|---|---|---|---|---|---|---|---|---|
| 总条件 | 2216E | 3 | 4 | 16 | 26 | 36 | 63 | 180 |
| R2A | 3 | 4 | 15 | 21 | 30 | 53 | 139 | |
| R | 4 | 5 | 16 | 25 | 35 | 62 | 152 | |
| SN | 4 | 5 | 15 | 27 | 44 | 73 | 203 | |
| 寡营养 | 2216E | 3 | 4 | 13 | 19 | 28 | 43 | 118 |
| R2A | 3 | 4 | 15 | 20 | 27 | 45 | 115 | |
| R | 4 | 5 | 13 | 20 | 28 | 40 | 91 | |
| SN | 3 | 4 | 12 | 19 | 30 | 43 | 104 | |
| 高盐(150‰ NaCl) | 2216E | 3 | 4 | 8 | 13 | 18 | 27 | 47 |
| R2A | 3 | 3 | 5 | 6 | 10 | 13 | 19 | |
| R | 3 | 3 | 8 | 13 | 18 | 24 | 47 | |
| SN | 4 | 5 | 10 | 15 | 20 | 32 | 73 | |
| 高温(50℃) | 2216E | 3 | 3 | 4 | 5 | 6 | 6 | 15 |
| R2A | 2 | 2 | 2 | 2 | 2 | 2 | 5 | |
| R | 3 | 4 | 5 | 6 | 11 | 12 | 14 | |
| SN | 3 | 5 | 5 | 5 | 6 | 9 | 26 |
表5 高温条件下4种培养基获得的物种情况Tab. 5 Species obtained by four selective media with the high temperature culture condition |
| 培养基 | 纲 | 种 |
|---|---|---|
| 5% 2216E | Actinomycetia | Streptomyces radiopugnans |
| Alphaproteobacteria | Acuticoccus kandeliae、Metabacillus endolithicus | |
| Bacilli | Bacillus altitudinis、Bacillus tequilensis、Cytobacillus pseudoceanisediminis | |
| 5% R2A | Alphaproteobacteria | Acuticoccus kandeliae |
| Actinomycetia | Streptomyces radiopugnans | |
| R | Actinomycetia | Streptomyces chumphonensis、Streptomyces genisteinicus、Streptomyces radiopugnans |
| Alphaproteobacteria | Microvirga flocculans、Pararhizobium herbae | |
| Bacilli | Bacillus altitudinis、Bacillus aequororis、Bacillus tequilensis、Halobacillus dabanensis、Metabacillus halosaccharovorans、Virgibacillus kapii、Mesobacillus boroniphilus | |
| SN | Actinomycetia | Streptomyces chumphonensis、Streptomyces ginkgonis、Streptomyces wuyuanensis |
| Alphaproteobacteria | Celeribacter ethanolicus、Lysobacter korlensis、Glycocaulis abyssi、Phenylobacterium terrae | |
| Bacilli | Bacillus mesophilus | |
| Gammaproteobacteria | Luteimonas pelagia |
| [1] |
韩敏敏, 李蜜, 刘昕明, 等, 2020. Khai岛和Pathiu岛珊瑚礁沉积物细菌多样性及细菌粗提物延缓秀丽隐杆线虫衰老活性研究[J]. 热带海洋学报, 39(5): 19-29.
|
| [2] |
何媛秋, 李存, 陈柔雯, 等, 2020. 不同培养条件对海洋沉积环境细菌的选择性分离[J]. 生物资源, 42(5): 540-548.
|
| [3] |
李存, 崔林青, 杨红强, 等, 2022. 三份南海岛礁珊瑚砂样品中可培养细菌多样性[J]. 热带海洋学报, 41(2): 149-158.
|
| [4] |
刘津江, 王淼, 樊敏, 等, 2022. 产脲酶微生物的筛选和应用研究进展[J]. 生物技术, 32(1): 107-113, 119.
|
| [5] |
柳鑫鹏, 臧淑英, 智刚, 等, 2022. 盐碱土耐盐碱细菌筛选及其植物促生能力研究[J]. 土壤通报, 53(3): 567-576.
|
| [6] |
邱路凡, 2023. 一株高效解钾菌解钾机理的探究及对土壤的改良[D]. 大庆: 东北石油大学.
|
| [7] |
王效昌, 马凯, 谢嘉慧, 等, 2023. 无机解磷菌对天鹅湖瀉湖沉积物内源磷释放的影响[J]. 农业资源与环境学报, 40(1): 76-85.
|
| [8] |
朱红惠, 黄力, 李文均, 等, 2021. 未/难培养微生物: 风正扬帆再起航, 共辉煌[J]. 微生物学报, 61(4):ⅰ–ⅱ.
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}